функционально активные поверхностные структуры толщиной в несколько молекулярных слоев, ограничивающие цитоплазму и большинство органелл клетки, а также образующие единую внутриклеточную систему канальцев, складок, замкнутых областей.
Биологические мембраны имеются во всех клетках. Их значение определяется важностью функций, которые они выполняют в процессе нормальной жизнедеятельности, а также многообразием заболеваний и патологических состояний, возникающих при различных нарушениях мембранных функций и проявляющихся практически на всех уровнях организации — от клетки и субклеточных систем до тканей, органов и организма в целом.
Мембранные структуры клетки представлены поверхностной (клеточной, или плазматической) и внутриклеточными (субклеточными) мембранами. Название внутриклеточных (субклеточных) мембран обычно зависит от названия ограничиваемых или образуемых ими структур. Так, различают митохондриальные, ядерные, лизосомные мембраны, мембраны пластинчатого комплекса аппарата Гольджи, эндоплазматического ретикулума, саркоплазматического ретикулума и др. (см. Клетка). Толщина биологических мембран — 7—10 нм, но их общая площадь очень велика, например, в печени крысы она составляет несколько сот квадратных метров.
Химический состав и строение биологических мембран. Состав М.б. зависит от их типа и функций, однако основными составляющими являются Липиды и Белки, а также Углеводы (небольшая, но чрезвычайно важная часть) и вода (более 20% общего веса).
Липиды. В составе М.б. обнаружены липиды трех классов: фосфолипиды, гликолипиды и стероиды. В мембранах животных клеток более 50% всех липидов составляют фосфолипиды — глицерофосфолипиды (фосфатидилхолин, фосфатидилэтаноламин, фосфатидилсерин, фосфатидилинозит) и сфингофосфолипиды (производные церамида, сфингомиелин). Гликолипиды представлены цереброзидами, сульфатидами и ганглиозидами, а стероиды — в основном холестерином (около 30%). В липидных компонентах М.б. содержатся разнообразные жирные кислоты, однако в мембранах животных клеток преобладают пальмитиновая, олеиновая и стеариновая кислоты. Основную структурную роль в биологических мембранах играют фосфолипиды. Они обладают выраженной способностью формировать двухслойные структуры (бислои) при смешивании с водой, что обусловлено химической структурой фосфолипидов, молекулы которых состоят из гидрофильной части — «головки» (остаток фосфорной кислоты и присоединенная к нему полярная группа, например холин) и гидрофобной части — «хвоста» (как правило, две жирно-кислотные цепи). В водной среде фосфолипиды бислоя расположены таким образом, что жирно-кислотные остатки обращены внутрь бислоя и, следовательно, изолированы от окружающей среды, а гидрофильные «головки» —наоборот, наружу. Липидный бислои представляет собой динамичную структуру: образующие его липиды могут вращаться, двигаться в латеральном направлении и даже переходить из слоя в слой (флип-флоп переход). Такое строение липидного бислоя легло в основу современных представлений о структуре М.б. и определяет некоторые важные свойства М.б., например способность служить барьером и не пропускать молекулы веществ, растворенных в воде (рис.). Нарушение структуры бислоя может привести к нарушению барьерной функции мембран.
Холестерин в составе М.б. играет роль модификатора бислоя, придавая ему определенную жесткость за счет увеличения плотности «упаковки» молекул фосфолипидов.
Гликолипиды несут разнообразные функции: отвечают за рецепцию некоторых биологически активных веществ, участвуют в дифференцировке ткани, определяют видовую специфичность.
Белки биологических мембран исключительно разнообразны. Молекулярная масса их в большинстве своем составляет 25 000 — 230 000.
Белки могут взаимодействовать с липидным бислоем за счет электростатических и (или) межмолекулярных сил. Они сравнительно легко могут быть удалены из мембраны. К такому типу белков относят цитохром с (молекулярная масса около 13 000), обнаруживаемый на наружной поверхности внутренней мембраны митохондрий.
Эти белки называются периферическими, или наружными. Для других белков, получивших название интегральных, или внутренних, характерно то, что одна или несколько полипептидных цепей оказываются погруженными в бислои или пересекают его, иногда не один раз (например, гликофорин, транспортные АТФ-азы, бактериородопсин). Часть белка, контактирующая с гидрофобной частью липидного бислоя, имеет спиральное строение и состоит из неполярных аминокислот, в силу чего между этими компонентами белков и липидов происходит гидрофобное взаимодействие. Полярные группы гидрофильных аминокислот непосредственно взаимодействуют с примембранными слоями, как с одной, так и с другой стороны бислоя. Молекулы белков, как и молекулы липидов, находятся в динамическом состоянии, для них также характерна вращательная, латеральная и вертикальная подвижность. Она является отражением не только их собственной структуры, но и функциональной активности. что в значительной степени определяется вязкостью липидного бислоя, которая, в свою очередь, зависит от состава липидов, относительного содержания и вида ненасыщенных жирно-кислотных цепей. Этим объясняется узкий температурный диапазон функциональной активности мембраносвязанных белков.
Белки мембран выполняют три основные функции: каталитическую (ферменты), рецепторную и структурную. Однако такое разграничение достаточно условно, и в ряде случаев один и тот же белок может выполнять и репепторную и ферментную функции (например, инсулин).
Число мембранных ферментов (Ферменты) в клетке достаточно велико, однако их распределение в различных типах М.б. неодинаково. Некоторые ферменты (маркерные) присутствуют только в мембранах определенного типа (например, Na, К-АТФ-аза, 5-нуклеотидаза, аденилатциклаза — в плазматической мембране; цитохром Р-450, НАДФН-дегидрогеназа, цитохром в5 — в мембранах эндоплазматического ретикулума; моноаминоксидаза — в наружной мембране митохондрий, а цитохром С-оксидаза, сукцинат-дегидрогеназа — во внутренней; кислая фосфатаза — в мембране лизосом).
Рецепторные белки, специфически связывая низкомолекулярные вещества (многие гормоны, медиаторы), обратимо меняют свою форму. Эти изменения запускают внутри клетки ответные химические реакции. Таким способом клетка принимает различные сигналы, поступающие из внешней среды.
К структурным белкам относят белки цитоскелета, прилегающие к цитоплазматической стороне клеточной мембраны. В комплексе с микротрубочками и микрофиламентами цитоскелета они обеспечивают противодействие клетки изменению ее объема и создают эластичность. В эту же группу включают ряд мембранных белков, функции которых не установлены.
Углеводы в биологических мембранах находятся в соединении с белками (гликопротеины) и липидами (гликолипиды). Углеводные цепи белков представляют собой олиго- или полисахаридные структуры, в состав которых входят глюкоза, галактоза, нейраминовая кислота, фукоза и манноза. Углеводные компоненты М.б. открываются в основном во внеклеточную среду, образуя на поверхности клеточных мембран множество ветвистых образований, являющихся фрагментами гликолипидов или гликопротеидов. Их функции связаны с контролем за межклеточным взаимодействием, поддержанием иммунного статуса клетки, обеспечением стабильности белковых молекул в М.б. Многие рецепторные белки содержат углеводные компоненты. Примером могут служить антигенные детерминанты групп крови, представленные гликолипидами и гликопротеинами.
Функции биологических мембран. Барьерная функция. Для клеток и субклеточных частиц М.б. служат механическим барьером, отделяющим их от внешнего пространства. Функционирование клетки часто сопряжено с наличием значительных механических градиентов на ее поверхности преимущественно вследствие осмотического и гидростатического давления. Основную нагрузку в этом случае несет клеточная стенка, главными структурными элементами которой у высших растений являются целлюлоза, пектин и экстепин, а у бактерий — муреин (сложный полисахарид-пептид). В клетках животных необходимость в жесткой оболочке отсутствует. Некоторую жесткость этим клеткам придают особые белковые структуры цитоплазмы, примыкающие к внутренней поверхности плазматической мембраны.
Перенос веществ через М.б. сопряжен с такими важнейшими биологическими явлениями, как внутриклеточный гомеостаз ионов, биоэлектрические потенциалы, возбуждение и проведение нервного импульса, запасание и трансформация энергии и т.п. (см. Биоэнергетика). Различают пассивный и активный транспорт (перенос) нейтральных молекул, воды и ионов через М.б. Пассивный транспорт не связан с затратами энергии, он осуществляется путем диффузии по концентрационным, электрическим или гидростатическим градиентам. Активный транспорт осуществляется против градиентов, связан с затратой энергии (преимущественно энергии гидролиза АТФ) и сопряжен с работой специализированных мембранных систем (мембранных насосов). Различают несколько видов транспорта. Если вещество транспортируется через мембрану независимо от наличия и переноса других соединений, то такой вид транспорта называют юнипортом. Если перенос одного вещества сопряжен с транспортом другого, то говорят о котранспорте, причем однонаправленный перенос называется симпортом, а противоположно направленный — антипортом. В особую группу выделяют перенос веществ путем экзо- и пиноцитоза.
Пассивный перенос может осуществляться путем простой диффузии через липидный бислои мембраны, а также через специализированные образования — каналы. Путем диффузии через мембрану проникают в клетку незаряженные молекулы, хорошо растворимые в липидах, в т.ч. многие яды и лекарственные средства, а также кислород и углекислый газ. Каналы представляют собой липопротеиновые структуры, пронизывающие мембраны. Они служат для переноса определенных ионов и могут находиться в открытом или закрытом состоянии. Проводимость канала зависит от мембранного потенциала, что играет важную роль в механизме генерации и проведения нервного импульса.
В ряде случаев перенос вещества совпадает с направлением градиента, но существенно превосходит по скорости простую диффузию. Этот процесс называют облегченной диффузией; он происходит с участием белков-переносчиков. Процесс облегченной диффузии не нуждается в энергии. Этим способом транспортируются сахара, аминокислоты, азотистые основания. Такой процесс происходит, например, при всасывании сахаров из просвета кишечника клетками эпителия.
Перенос молекул и ионов против электрохимического градиента (активный транспорт) связан со значительными затратами энергии. Часто градиенты достигают больших величин. например, концентрационный градиент водородных ионов на плазматической мембране клеток слизистой оболочки желудка составляет 106, градиент концентрации ионов кальция на мембране саркоплазматического ретикулума — 104, при этом потоки ионов против градиента значительны. В результате затраты энергии на транспортные процессы достигают, например, у человека, более 1/3 всей энергии метаболизма. В плазматических мембранах клеток различных органов обнаружены системы активного транспорта ионов натрия и калия — натриевый насос. Эта система перекачивает натрий из клетки и калий в клетку (антипорт) против их электрохимических градиентов. Перенос ионов осуществляется основным компонентом натриевого насоса — Na+, К+-зависимой АТФ-азой за счет гидролиза АТФ. На каждую гидролизующуюся молекулу АТФ транспортируется три иона натрия и два иона калия. Существуют два типа Са2+-АТФ-аз. Одна из них обеспечивает выброс ионов кальция из клетки в межклеточную среду, другая — аккумуляцию кальция из клеточного содержимого во внутриклеточное депо. Обе системы способны создавать значительный градиент иона кальция. К+, Н+-АТФ-аза обнаружена в слизистой оболочке желудка и кишечника. Она способна транспортировать Н+ через мембрану везикул слизистой оболочки при гидролизе АТФ. В микросомах слизистой оболочки желудка лягушки найдена аниончувствительная АТФ-аза, способная при гидролизе АТФ осуществлять антипорт бикарбоната и хлорида.
Изложенные механизмы транспорта различных веществ через клеточные мембраны имеют место и в случае их транспорта через эпителий ряда органов (кишечника, почек, легких), который осуществляется через слой клеток (монослой в кишечнике и нефронах), а не через единичную клеточную мембрану. Такой транспорт называют трансцеллюлярным, или трансэпителиальным. Характерной особенностью клеток, например эпителиоцитов кишечника и канальцев нефронов, является то, что апикальная и базальная их мембраны различаются по проницаемости, величине мембранного потенциала и транспортной функции.
Способность генерировать биоэлектрические потенциалы и проводить возбуждение. Возникновение биоэлектрических потенциалов связано с особенностями строения биологических мембран и с деятельностью их транспортных систем, создающих неравномерное распределение ионов по обе стороны мембраны (см. Биоэлектрические потенциалы, Возбуждение).
Процессы трансформации и запасания энергии протекают в специализированных М.б. и занимают центральное место в энергетическом обеспечении живых систем. Два основных процесса энергообразования — фотосинтез и тканевое дыхание — локализованы в мембранах внутриклеточных органелл высших организмов, а у бактерий — в клеточной (плазматической) мембране (см. Дыхание тканевое). Фотосинтезирующие мембраны преобразуют энергию света в энергию химических соединений, запасая ее в форме сахаров — основного химического источника энергии для гетеротрофных организмов. При дыхании энергия органических субстратов освобождается в процессе переноса электронов по цепи окислительно-восстановительных переносчиков и утилизируется в процессе фосфорилирования АДФ неорганическим фосфатом с образованием АТФ. Мембраны, осуществляющие фосфорилирование, сопряженное с дыханием, называют сопрягающими (внутренние мембраны митохондрий, клеточные мембраны некоторых аэробных бактерий, мембраны хроматофоров фотосинтезирующих бактерий).
Метаболические функции мембран определяются двумя факторами: во-первых, связью большого числа ферментов и ферментативных систем с мембранами, во-вторых, способностью мембран физически разделять клетку на отдельные отсеки, отграничивая друг от друга метаболические процессы, протекающие в них. Метаболические системы не остаются при этом полностью изолированными. В мембранах, разделяющих клетку, имеются специальные системы, обеспечивающие избирательное поступление субстратов, выделение продуктов, а также движение соединений, обладающих регуляторным действием.
Клеточная рецепция и межклеточные взаимодействия. Под этой формулировкой объединен весьма обширный и разнообразный набор важных функций клеточных мембран, определяющих взаимодействие клетки с окружающей средой и формирование многоклеточного организма как единого целого. Молекулярно-мембранные аспекты клеточной рецепции и межклеточных взаимодействий касаются прежде всего иммунных реакций, гормонального контроля роста и метаболизма, закономерностей эмбрионального развития.
Нарушения структуры и функции биологических мембран. Разнообразие типов М.б., их полифункциональность и высокая чувствительность к внешним условиям порождают необыкновенное разнообразие структурно-функциональных нарушений мембран, возникающих при многих неблагоприятных воздействиях и сопряженных с огромным числом конкретных заболеваний организма как целого. Все это разнообразие нарушений достаточно условно можно подразделить на транспортные, функционально-метаболические и структурные. В общем виде охарактеризовать последовательность возникновения этих нарушений не представляется возможным, и в каждом конкретном случае требуется детальный анализ для выяснения первичного звена в цепи развития структурно-функциональных нарушений мембран. Нарушение транспортных функций мембран, в частности увеличение проницаемости мембран, — общеизвестный универсальный признак повреждения клетки. Нарушением транспортных функций (например, у человека) обусловлено более 20 так называемых транспортных болезней, среди которых почечная глюкозурия, цистинурия, нарушение всасывания глюкозы, галактозы и витамина В12, наследственный сфероцитоз и др. Среди функционально-метаболических нарушений М.б. центральными являются изменения процессов биосинтеза, а также многообразные отклонения в энергообеспечении живых систем. В наиболее общем виде следствием этих процессов является нарушение состава и физико-химических свойств мембран, выпадение отдельных звеньев метаболизма и его извращение, а также снижение уровня жизненно важных энергозависимых процессов (активного транспорта ионов, процессов сопряженного транспорта, функционирования сократительных систем и т.д.). Повреждения ультраструктурной организации М.б. выражаются в чрезмерном везикулообразовании, увеличении поверхности плазматических мембран за счет образования пузырей и отростков, слиянии разнородных клеточных мембран, образовании микропор и локальных структурных дефектов.
Библиогр.: Биологические мембраны, под ред. Д.С. Парсонса, пер. с англ., М., 1978; Болдырев А.А. Введение в биохимию мембран, М., 1986, библиогр.; Конев С.В. и Мажуль В.М. Межклеточные контакты. Минск, 1977; Кульберг А.Я. Рецепторы клеточных мембран, М., 1987, библиогр.; Маленков А.Г. и Чуич Г.А. Межклеточные контакты и реакции ткани, М., 1979; Сим Э. Биохимия мембран, пер. с англ., М., 1985, библиогр.; Финеан Дж., Колмэн Р. и Митчелл Р. Мембраны и их функции в клетке, пер. с англ., М., 1977, библиогр.
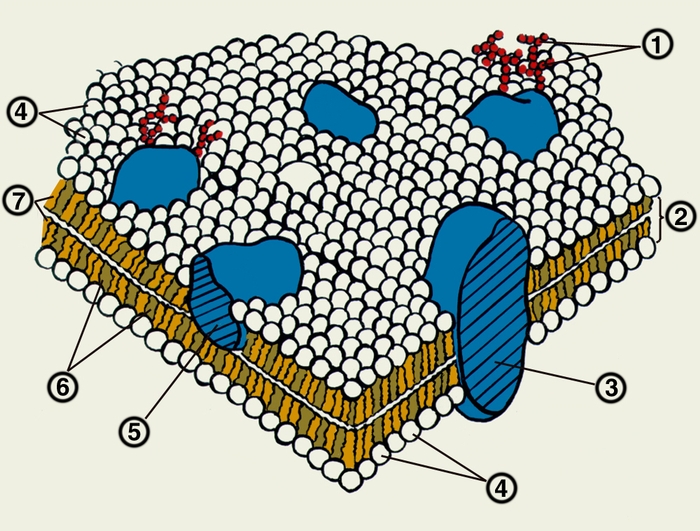 холестерин; 7 — жирнокислотные «хвосты» фосфолипидов">
холестерин; 7 — жирнокислотные «хвосты» фосфолипидов">Схема строения биологической мембраны: 1 — углеводные фрагменты гликопротеидов; 2 — липидный бислой; 3 — интегральный белок; 4 — «головки» фосфолипидов; 5 — периферический белок; 6 — холестерин; 7 — жирнокислотные «хвосты» фосфолипидов.
1. Малая медицинская энциклопедия. — М.: Медицинская энциклопедия. 1991—96 гг. 2. Первая медицинская помощь. — М.: Большая Российская Энциклопедия. 1994 г. 3. Энциклопедический словарь медицинских терминов. — М.: Советская энциклопедия. — 1982—1984 гг.