мейоз, деление созревания
Двухступенчатое деление клеток, приводящее к образованию из диплоидных клеток гаплоидных, что является основным этапом гаметогенеза; выделяют 3 типа М.: зиготный, или начальный (у многих грибов и водорослей) - происходит сразу после оплодотворения и приводит к образованию гаплоидного таллома или мицелия, гаметный, или конечный (у всех многоклеточных животных и у некоторых низших растений) - происходит в половых органах и приводит к образованию гамет, споровый, или промежуточный (у высших растений) - происходит перед цветением и приводит к образованию гаплоидного гаметофита, у простейших встречаются все 3 типа М.; М. включает два деления, разделенных интеркинезом interkinesis (но не всегда обязательным), - I деление характеризуется очень длинной, дифференцированной на стадии профазой, во II профаза и метафаза могут выпадать; удвоение ДНК происходит только перед I делением М.; однако прежняя точка зрения о том, что в I делении расходятся гомологичные хромосомы, а во II - хроматиды (т.е. I - редукционное деление, а II - эквационное) не подтверждается: в I делении расходятся либо хромосомы, либо хроматиды, а во II - наоборот; М. был открыт У.Флеммингом у животных в 1882 и Э.Страсбургером у растений в 1888.
* * *
Мейоз — деление клеточного ядра, предшествующее образованию половых клеток и связанное с уменьшением (редукцией) числа хромосом, свойственного соматической клетке, в 2 раза. Различают 3 типа М.:
а) начальный, или зиготный, — происходит сразу после оплодотворения, приводя к образованию гаплоидного таллома или мицелия (у многих грибов и водорослей);
б) конечный, или гаметный, — происходит в половых органах и приводит к образованию гамет (у всех многоклеточных животных и у некоторых низших растений);
в) споровый, или промежуточный, — происходит перед цветением и приводит к образованию гаплоидного гаметофита (у высших растений).
У простейших выявлены все 3 типа М. Гаметный (конечный) тип М. состоит из двух следующих одно за другим делений: I, которое включает в себя очень длинную, состоящую из нескольких стадий профазу и метафазу, и II, в котором могут выпадать профаза и метафаза. Профазы I деления подразделяется на 5 последовательных стадий: лептотену (лептонему), зиготену (зигонему), пахитену (пахинему), диплотену (диплонему) и диакинез. В течение лептотены хромосомы имеют вид тонких нитей с ясно различимыми хромомерами. Все хромосомы часто ориентированы одним или обоими концами и контактируют с одним участком ядерной мембраны, образуя конфигурацию «букета». Каждая хромосома состоит из 2 хроматид, однако это остается неразличимым до пахитены (репликация ДНК и удвоение ее диплоидного количества происходят до наступления лептотены). В диплоидных соматических клетках (2N) хромосомы присутствуют в виде N пар и каждая хромосома является репликантом одной из родительских хромосом самца и самки в данной зиготе. В ядрах соматических клеток большинства организмов гомологичные хромосомы не образуют пары. В М. в течение стадии зиготены происходит синапсис гомологичных хромосом: образование пар начинается в ряде точек и продолжается до полного завершения конъюгации (см. Конъюгация хромосом). Этот процесс сопровождается формированием синаптонемального комплекса. Когда синапсис заканчивается, реальное число хромосомных нитей равно половине того числа, которое было ранее, и они различимы в ядре как биваленты, а не единичные хромосомы. На стадии пахитены каждая парная хромосома разделяется на две сестринские хроматиды (за исключением центромеры). В результате продольного деления каждой гомологичной хромосомы на 2 хроматиды в ядре образуется N групп из 4 хроматид, лежащих параллельно друг другу, называемых тетрадами. Происходит локализованный разрыв с последующим обменом участками между несестринскими хроматидами — кроссинговер (см). Этот процесс сопровождается синтезом конститутивной ДНК в количестве меньшем, чем 1% от всего его количества в ядре. Обмен между гомологичными хромосомами приводит к образованию кроссоверных хроматид (см. Кроссоверы), содержащих генетический материал и отцовского, и материнского происхождения. На протяжении стадии диплотены одна из пар сестринских хроматид в каждой из тетрад начинает отделяться от др. пары. Однако хроматиды не разделяются в том месте, где имел место обмен, — в таких районах частичного перекрытия хроматиды образуют крестообразную структуру, называемую хиазмой. Число хиазм зависит от вида хромосомы и прямо пропорционально ее длине. Затем происходит терминализация хиазм, которая продолжается в течение диакинеза до тех пор, пока все хиазмы не достигнут концов тетрад и гомологи смогут разделиться во время анафазы. В диакинезе хромосомы плотно спирализуются, укорачиваются и утолщаются, образуя группу компактных тетрад, хорошо упакованных в ядре, чаще всего около его мембраны. Терминализация полностью завершается и исчезает ядрышко. Во время I деления исчезает оболочка ядра и тетрады располагаются в области экватора, где находится веретено деления. Хроматиды тетрад разъединяются т. обр., что происходит разделение материнского и отцовского генетического материала, за исключением дистального участка, где произошел кроссинговер. Во время I деления образуются 2 вторичных гаметоцита, которые содержат диады, окруженные ядерной оболочкой. II деление начинается после короткой интерфазы, в течение которой хромосомы не спирализуются. Ядерная мембрана исчезает, и диады располагаются на метафазной пластинке. Хроматиды каждой диады эквивалентны друг другу (за исключением дистальных участков с точками, претерпевшими кроссинговер), центромера делится, и каждая хромосома получает возможность уйти в отдельную клетку. У животных во время II деления образуются 4 сперматиды или оотиды с гаплоидным набором хромосом, окруженные ядерной мембраной. М., т. обр., обеспечивает механизм, посредством которого происходит обмен генетическим материалом между гомологичными хромосомами и каждая гамета получает по одной из пары хромосом. В последнее время появляются сообщения о том, что прежняя точка зрения о расхождении гомологичных хромосом в I делении М. (редукционное деление), а хроматид — во II (эквационное) не подтверждается: в I делении расходятся либо хромосомы, либо хроматиды, а во II — наоборот. М. открыт У.Флемингом у животных в 1882 г. и Э.Стасбургером у растений в 1888 г. 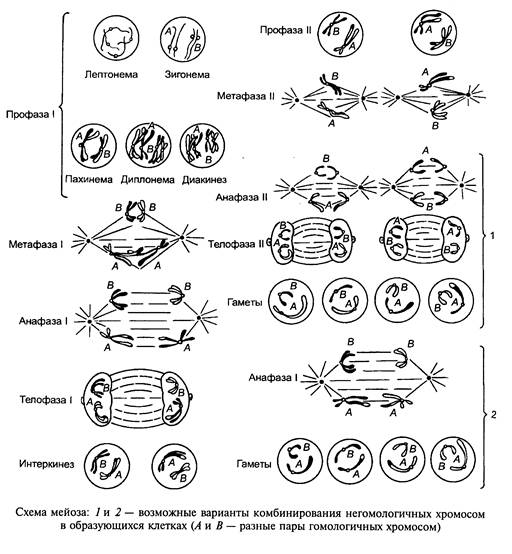
Англо-русский толковый словарь генетических терминов. — М.: Изд-во ВНИРО. Арефьев В.А., Лисовенко Л.А., науч. ред. Л.И.Патрушев. 1995.