элементарная живая система, способная к самостоятельному существованию, самовоспроизведению и развитию; основа строения и жизнедеятельности всех животных и растений. К. существуют и как самостоятельные организмы (см. Простейшие), и в составе многоклеточных организмов (тканевые К.). Термин «К.» предложен английским микроскопистом Р. Гуком (1665). К. — предмет изучения особого раздела биологии — цитологии (См. Цитология). Систематическое изучение К. началось лишь в 19 в. Одним из крупнейших научных обобщений того времени была Клеточная теория, утверждавшая единство строения всей живой природы. Изучение жизни на клеточном уровне лежит в основе современных биологических исследований.
В строении и функциях каждой К. обнаруживаются признаки, общие для всех К., что отражает единство их происхождения из первичных органических комплексов. Частные особенности различных К. — результат их специализации в процессе эволюции. Так, все К. сходно регулируют обмен веществ, удваивают и используют свой наследственный материал, получают и утилизируют энергию. В то же время разные одноклеточные организмы (амёбы, инфузории и т.д.) сильно различаются размерами, формой, поведением. Не менее резко различаются К. многоклеточных организмов. Так, у человека имеются лимфоидные К. — небольшие (диаметром около 10 мкм) округлые К., участвующие в иммунологических реакциях, и нервные К., часть которых имеет отростки длиной более метра; эти К. осуществляют основные регуляторные функции в организме.
Методы исследования. Первым цитологическим методом была микроскопия живых К. Современные варианты прижизненной (витальной) световой микроскопии — фазово-контрастная, люминесцентная, интерференционная и др. (см. Микроскоп) — позволяют изучать форму К. и общее строение некоторых её структур, движение К. и их деление. Детали строения К. обнаруживаются лишь после специального контрастирования, что достигается окраской убитой К. Новый этап изучения структуры К. — электронная микроскопия, дающая значительно большее разрешение структур К. по сравнению со световой микроскопией (см. Разрешающая способность оптических приборов). Химический состав К. изучается цито- и гистохимическими методами, позволяющими выяснить локализацию и концентрацию веществ в клеточных структурах, интенсивность синтеза веществ и их перемещение в К. (см. Гистохимия). Цитофизиологические методы позволяют изучать функции К., например возбуждение, секрецию. См. также Авторадиография, Микроскопическая техника, Цитофотометрия.
Общие свойства клеток. В каждой К. различают две основные части — Ядро и цитоплазму (См. Цитоплазма), в которых, в свою очередь, можно выделить структуры, различающиеся по форме, размерам, внутреннему строению, химическим свойствам и функциям. Одни из них — так называемые органоиды — жизненно необходимы К. и обнаруживаются во всех К. Другие — продукты активности К., представляющие временные образования. В специализированных структурах осуществляется разделение различных биохимических функций, что способствует осуществлению в одной и той же К. разнородных процессов, включающих синтез и распад многих веществ.
В ядерных органоидах — хромосомах (См. Хромосомы), в их основном компоненте — ДНК, хранится генетическая информация о строении белков, свойственных организму определённого вида (см. Ген, Генетический код). Другое важнейшее свойство ДНК — способность к самовоспроизведению, что обеспечивает как стабильность наследственной информации, так и её непрерывность — передачу следующим поколениям. На ограниченных участках ДНК, охватывающих несколько генов, как на матрицах, синтезируются рибонуклеиновые кислоты — непосредственные участники синтеза белка. Перенос (Транскрипция) кода ДНК происходит при синтезе информационных РНК (и-РНК). Синтез белка представляется как считывание информации с матрицы РНК. В этом процессе, называемом трансляцией (См. Трансляция), принимают участие транспортные РНК (т-РНК) и специальные органоиды — Рибосомы, образующиеся в ядрышке (См. Ядрышко). Размеры ядрышка определяются главным образом потребностью К. в рибосомах; поэтому особенно велико оно в К., интенсивно синтезирующих белок. Синтез белка — конечный итог реализации функций хромосом — осуществляется главным образом в цитоплазме. Белки — ферменты, детали структур и регуляторы разных процессов, включая и транскрипцию — определяют в конечном счёте все стороны жизни К., позволяя К. сохранять свою индивидуальность, несмотря на постоянно меняющееся окружение. Если в бактериальной К. синтезируется около 1000 разных белков, то почти в каждой из К. человека — свыше 10000. Таким образом, разнообразие внутриклеточных процессов в ходе эволюции организмов существенно возрастает. Оболочка ядра, отделяющая его содержимое от цитоплазмы, состоит из двух мембран, пронизанных порами — специализированных участков для транспорта некоторых соединений из ядра в цитоплазму и обратно. Другие вещества проходят через мембраны путём диффузии или активного транспорта, требующего затрат энергии. Многие процессы происходят в цитоплазме К. при участии мембран эндоплазматической сети (См. Эндоплазматическая сеть) — основной синтезирующей системы К., а также Гольджи комплекса и митохондрий (См. Митохондрии). Отличия мембран разных органоидов определяются свойствами образующих их белков и липидов. К некоторым мембранам эндоплазматической сети прикреплены рибосомы; здесь происходит интенсивный синтез белка. Такая гранулярная эндоплазматическая сеть особенно развита в К., секретирующих или интенсивно обновляющих белок, например у человека в К. печени, поджелудочной железы, нервных К. В состав других биологических мембран (См. Биологические мембраны), лишённых рибосом (гладкоконтурная сеть), входят ферменты, участвующие в синтезе углеводно-белковых и липидных комплексов. В каналах эндоплазматической сети могут временно накапливаться продукты деятельности К.; в некоторых К. по каналам происходит направленный транспорт веществ. Перед выведением из К. вещества концентрируются в пластинчатом комплексе (комплексе Гольджи). Здесь обособляются различные включения К., например секреторные или пигментные гранулы, образуются Лизосомы — пузырьки, содержащие гидролитические ферменты и участвующие во внутриклеточном переваривании многих веществ. Система окруженных мембранами каналов, вакуолей и пузырьков представляет одно целое. Так, эндоплазматическая сеть может без перерыва переходить в мембраны, окружающие ядро, соединяться с цитоплазматической мембраной, формировать комплекс Гольджи. Однако связи эти нестабильны. Нередко, а во многих К. обычно разные мембранные структуры разобщены и обмениваются веществами через гиалоплазму (См. Гиалоплазма). Энергетика К. во многом зависит от работы митохондрий. Число их колеблется в К. разного типа от десятков до тысяч. Например, в печёночной К. человека около 2 тыс. митохондрий; их общий объём не менее 1/5 объёма К. Внешняя мембрана митохондрии отграничивает её от цитоплазмы, на внутренней — происходят основные энергетические превращения веществ, в результате которых образуется соединение, богатое энергией, — аденозинтрифосфорная кислота (АТФ) — универсальный переносчик энергии в К. Митохондрии содержат ДНК и способны к самовоспроизведению; однако автономность митохондрий относительна, их репродукция и деятельность зависят от ядра. За счет энергии АТФ в К. осуществляются различные синтезы, транспорт и выделение веществ, механическая работа, регуляция процессов и т.д. В делении К. и иногда в их движении участвуют структуры, имеющие вид трубочек субмикроскопических размеров. «Сборка» таких структур и их функционирование зависят от центриолей (См. Центриоли), при участии которых организуется Веретено деления клетки, с чем связано перемещение хромосом и ориентация оси деления К. Базальные тельца — производные центриолей — необходимы для построения и нормальной работы жгутиков и ресничек — локомоторных и чувствительных образований К., строение которых у простейших и в различных К. многоклеточных однотипно.
От внеклеточной среды К. отделена плазматической мембраной, через которую происходит поступление ионов и молекул в К. и выделение их из К. Отношение поверхности К. к ее объему уменьшается с увеличением объема, и чем крупнее К., тем более затруднены ее связи с внешней средой. Величина К. не может быть особенно большой. Для живых К. характерен Активный транспорт ионов, требующий затраты энергии, специальных ферментов и, возможно, переносчиков. Благодаря активному и избирательному переносу в К. одних ионов и непрерывному удалению из нее других создается разность концентраций ионов в К. и окружающей среде. Этот эффект может быть обусловлен и связыванием ионов компонентами К. Многие ионы необходимы как активаторы внутриклеточных синтезов и как стабилизаторы структуры органоидов. Обратимые изменения соотношения ионов в К. и среде лежат в основе биоэлектрической активности К. — одного из важных факторов передачи сигналов от одной К. к другой (см. Биоэлектрические потенциалы). Образуя впячивания, которые затем замыкаются и отделяются в виде пузырьков внутрь К., плазматическая мембрана способна захватывать растворы крупных молекул (Пиноцитоз) или даже отдельные частицы величиной в несколько мкм (Фагоцитоз). Так осуществляется питание некоторых К., перенос веществ через К., захват бактерий фагоцитами. Со свойствами плазматической мембраны связаны и силы сцепления, удерживающие во многих случаях К. друг около друга, например в покровах тела или внутренних органах. Сцепление и связь К. обеспечиваются химическим взаимодействием мембран и специальными структурами мембраны — десмосомами (См. Десмосомы).
Рассмотренная в общей форме схема строения К. свойственна в основных чертах как животным, так и растительным К.. Но есть и существенные отличия в особенностях метаболизма и строения растительных К. от животных.
Клетки растений. Поверх плазматической мембраны растительные К. покрыты, как правило, твёрдой внешней оболочкой (она может отсутствовать лишь у половых К.), состоящей у большинства растений главным образом из полисахаридов: целлюлозы, пектиновых веществ и гемицеллюлоз, а у грибов и некоторых водорослей — из хитина. Оболочки снабжены порами, через которые с помощью выростов цитоплазмы соседние К. связаны друг с другом. Состав и строение оболочки меняются по мере роста и развития К. Часто у К., прекративших рост, оболочка пропитывается лигнином, кремнезёмом или др. веществом, которое делает её более прочной. Оболочки К. определяют механические свойства растения. К. некоторых растительных тканей отличаются особенно толстыми и прочными стенками (см. Древесина), сохраняющими свои скелетные функции после гибели К. Дифференцированные растительные К. имеют несколько вакуолей (См. Вакуоли) или одну центральную вакуоль, занимающую обычно большую часть объёма К. Содержимое вакуолей — раствор различных солей, углеводов, органических кислот, алкалоидов, аминокислот, белков, а также запас воды. В вакуолях могут откладываться питательные вещества. В цитоплазме растительной К. имеются специальные органоиды — Пластиды; лейкопласты (в них часто откладывается крахмал), хлоропласты (содержат преимущественно хлорофилл и осуществляют Фотосинтез) и хромопласты (содержат пигменты из группы каротиноидов). Пластиды, как и митохондрии, способны к самовоспроизведению. Комплекс Гольджи в растительной К. представлен рассеянными по цитоплазме диктиосомами (См. Диктиосомы).
Одноклеточные организмы. В строении и функциях одноклеточных, или простейших, черты, свойственные любой К., сочетаются с признаками самостоятельных организмов. Так, у простейших такой же набор органоидов, как и у К. многоклеточных; идентично и ультрастроение их органоидов; при делении простейших в них обнаруживаются типичные хромосомы. Однако приспособление простейших к разным средам обитания (водной или наземной, к свободному или паразитическому существованию) обусловило существенное разнообразие их строения и физиологии. Многие простейшие (жгутиковые, инфузории) обладают сложным двигательным аппаратом и имеют органеллы, связанные с захватом пищи и пищеварением. Изучение простейших представляет большой интерес для выяснения филогенетических возможностей К.: эволюционные изменения организма протекают у них на клеточном уровне. В отличие от простейших и К. многоклеточных организмов, бактерии, синезеленые водоросли, актиномицеты не имеют оформленного ядра и хромосом. Их генетический аппарат, называется нуклеоидом, представлен нитями ДНК и не окружен оболочкой. Еще более отличаются от К. многоклеточных организмов и от простейших Вирусы, у которых отсутствуют основные, необходимые для обмена веществ ферменты. Поэтому вирусы могут расти и размножаться, лишь проникая в К. и используя их ферментные системы.
Специальные функции клеток. В процессе эволюции многоклеточных возникло разделение функций между К., что привело к расширению возможностей приспособления животных и растений к меняющимся условиям среды. Закрепившиеся наследственно различия в форме К., их размерах и некоторых сторонах метаболизма реализуются в процессе индивидуального развития организма. Основное проявление развития — Дифференцировка К., их структурная и функциональная специализация. Дифференцированные К. имеют такой же набор хромосом, как и оплодотворенная яйцеклетка. Это доказывается пересадкой ядра дифференцированной К. в предварительно лишенную ядра яйцеклетку, после чего может развиваться полноценный организм. Таким образом, различия между дифференцированными К., по-видимому, обусловливаются разными соотношениями активных и неактивных генов, каждый из которых кодирует биосинтез определённого белка. Судя по составу белков, в дифференцированных К. активна (способна к транскрипции) лишь небольшая часть (порядка 10%) генов, свойственных К. данного вида организмов. Среди них лишь немногие ответственны за специальную функцию К., а остальные обеспечивают общеклеточные функции. Так, в мышечных К. активны гены, кодирующие структуру сократимых белков, в эритроидных К. — гены, кодирующие биосинтез гемоглобина, и т.д. Однако в каждой К. должны быть активны гены, определяющие биосинтез веществ и структур, необходимых для всех К., например ферментов, участвующих в энергетических превращениях веществ. В процессе специализации К. отдельные общеклеточные функции их могут развиваться особенно сильно. Так, в железистых К. более всего выражена синтетическая активность, мышечные — наиболее сократимы, нервные — наиболее возбудимы. В узкоспециализированных К. обнаруживаются структуры, характерные лишь для этих К. (например, у животных — миофибриллы мышц, тонофибриллы и реснички некоторых покровных К., нейрофибриллы нервных К., жгутики у простейших или у сперматозоидов многоклеточных организмов). Иногда специализация сопровождается утратой некоторых свойств (например, нервные К. утрачивают способность к размножению; ядра К. кишечного эпителия млекопитающих не могут в зрелом состоянии синтезировать РНК; зрелые эритроциты млекопитающих лишены ядра). Выполнение важных для организма функций включает иногда гибель К. Так, К. эпидермиса кожи постепенно ороговевают и гибнут, но остаются некоторое время в пласте, предохраняя подлежащие ткани от повреждения и инфекции. В сальных железах К. постепенно превращаются в капли жира, который используется организмом или выделяется. Для выполнения некоторых тканевых функций К. образуют неклеточные структуры. Основные пути их образования — секреция или превращения компонентов цитоплазмы. Так, значительная по объёму часть подкожной клетчатки, хряща и кости составляет межуточное вещество — производное К. соединительной ткани. К. крови обитают в жидкой среде (плазме крови), содержащей белки, сахара и др. вещества, вырабатываемые разными К. организма. К. эпителия, образующие пласт, окружены тонкой прослойкой диффузно распределённых веществ, главным образом гликопротеидов (так называемый цемент, или надмембранный компонент). Внешние покровы членистоногих и раковины моллюсков — также продукты выделения К. Взаимодействие специализированных К. — необходимое условие жизни организма и нередко самих этих К. (см. Гистология). Лишённые связей друг с другом, например в культуре, К. быстро утрачивают особенности присущих им специальных функций.
Деление клеток. В основе способности К. к самовоспроизведению лежат уникальное свойство ДНК самокопироваться и строго равноценное деление репродуцированных хромосом в процессе Митоза. В результате деления образуются две К., идентичные исходной по генетическим свойствам и с обновленным составом ядра и цитоплазмы. Процессы самовоспроизведения хромосом, их деления, образования двух ядер и деления цитоплазмы разделены во времени, составляя в совокупности Митотический цикл К. В случае, если после деления К. начинает готовиться к следующему делению, митотический цикл совпадает с жизненным циклом К. Однако во многих случаях после деления (а иногда перед ним) К. выходят из митотического цикла, дифференцируются и выполняют в организме ту или иную специальную функцию. Состав таких К. может обновляться за счёт делений малодифференцированных К. В некоторых тканях и дифференцированные К. способны повторно входить в митотический цикл. В нервной ткани дифференцированные К. не делятся; многие из них живут так же долго, как организм в целом, то есть у человека — несколько десятков лет. При этом ядра нервных К. не утрачивают способности к делению: будучи пересажены в цитоплазму раковых К., ядра нейронов синтезируют ДНК и делятся. Опыты с клетками-гибридами показывают влияние цитоплазмы на проявление ядерных функций. Неполноценная подготовка к делению предотвращает митоз или искажает его течение. Так, в некоторых случаях не происходит деления цитоплазмы и образуется двуядерная К. Многократное деление ядер в неделящейся К. приводит к появлению многоядерных К. или сложных надклеточных структур (симпластов), например в поперечнополосатых мышцах. Иногда репродукция К. ограничивается воспроизведением хромосом, и образуется полиплоидная К., имеющая удвоенный (сравнительно с исходной К.) набор хромосом. Полиплоидизация приводит к усилению синтетической активности, увеличению размеров и массы К.
Обновление клеток. Для длительной работы каждой К. необходимо восстановление изнашиваемых структур, как и ликвидация повреждений К., вызванных внешними воздействиями. Восстановительные процессы, характерные для всех К., связаны с изменениями проницаемости плазматической мембраны и сопровождаются усилением внутриклеточных синтезов, в первую очередь синтеза белка. Во многих тканях стимуляция восстановительных процессов приводит к репродукции генетического аппарата и делению К.; это свойственно, например покровам или кроветворной системе. Процессы внутриклеточного обновления в этих тканях выражены слабо, их К. живут сравнительно недолго (например, К. кишечного покрова млекопитающих — всего несколько суток). Максимальной выраженности внутриклеточные восстановительные процессы достигают в неделящихся или слабоделящихся клеточных популяциях, например в нервных К. Показателем совершенства процессов внутреннего обновления К. является длительность их жизни; для многих нервных К. она совпадает с продолжительностью жизни всего организма.
Мутации. Обычно процесс воспроизведения ДНК происходит без отклонений, и генетический код остаётся постоянным, что обеспечивает синтез одного и того же набора белков в огромном числе клеточных поколений. Однако в редких случаях может произойти мутация (См. Мутации) — частичное изменение структуры гена. Конечный её эффект — изменение свойств белков, кодируемых мутантными генами. Если при этом затрагиваются важные ферментные системы, свойства К., а иногда и всего организма существенно изменяются. Так, мутация одного из генов, контролирующих синтез гемоглобина, приводит к тяжелому заболеванию — анемии (См. Анемия). Естественный отбор полезных мутаций — важный механизм эволюции.
Регуляция функций клеток. Основной механизм регуляции внутриклеточных процессов связан с различными влияниями на ферменты — высоко специфичные катализаторы биохимических реакций. Регуляция может осуществляться на генетическом уровне, когда определяется состав ферментов или количество того или иного фермента в К. В последнем случае регуляция может происходить и на уровне трансляции. Другой тип регуляции — воздействие на сам фермент, в результате чего может происходить как торможение, так и стимуляция его активности. Структурный уровень регуляции — влияние на сборку клеточных структур: мембран, рибосом и т.д. Конкретными регуляторами внутриклеточных процессов могут быть нервные влияния, гормоны, специальные вещества, вырабатываемые внутри К. либо окружающими К. (особенно белки), или же сами продукты реакций. В последнем случае воздействие осуществляется по принципу обратной связи, когда продукт реакции влияет на активность фермента — катализатора этой реакции. Регуляция может осуществляться через транспорт предшественников и ионов, влияния на матричный синтез (РНК, полисомы, ферменты синтеза), изменение формы регулируемого фермента.
Организация и регуляция функций К. на молекулярном уровне определяют такие свойства живых систем, как пространственная компактность и энергетическая экономичность. Важное свойство многоклеточных организмов — надёжность — во многом зависит от множественности (взаимозаменяемости) К. каждого функционального типа, а также от возможности их замены в результате размножения К. и обновления компонентов каждой К.
В медицине используются воздействия на К. для лечения и предупреждения заболеваний. Многие лекарственные вещества изменяют активность определенных К. Так, наркотики, транквилизаторы и болеутоляющие вещества снижают интенсивность деятельности нервных К., а стимуляторы её усиливают. Некоторые вещества стимулируют сокращение мышечных К. сосудов, другие — матки или сердца. Специальные воздействия на делящиеся К. осуществляются при использовании радиации или цитостатических веществ, блокирующих деление К. Иммунизация стимулирует деятельность лимфоидных К., вырабатывающих антитела к чужеродным белкам, предупреждая тем самым многие заболевания.
Лит.: Кольцов Н. К., Организация клетки, М. — Л., 1936; Вильсон Э., Клетка и её роль в развитии и наследственности, пер. с англ., т. 1—2, М. — Л., 1936—1940; Насонов Д. Н. и Александров В. Я., Реакция живого вещества на внешние воздействия, М. — Л., 1940; Кедровский Б. В., Цитология белковых синтезов в животной клетке, М., 1959; Мэзия Д., Митоз и физиология клеточного деления, пер. с англ., М., 1963; Руководство по цитологии, т. 1—2, М. — Л., 1965—66; Бродский В. Я., Трофика клетки, М., 1966; Живая клетка, [Сб. ст.], пер. с англ., М., 1966; Де Робертис Э., Новинский В., Саэс Ф., Биология клетки, пер. с англ., М., 1967; Васильев Ю. М. и Маленков А. Г., Клеточная поверхность и реакции клеток, Л., 1968; Алов И. А., Брауде А. И., Аспиз М. Е., Основы функциональной морфологии клетки, 2 изд., М., 1969; Лёви А., Сикевиц Ф., Структура и функции клетки, пер. с англ., М., 1971; Handbook of molecular cytology, ed. A. Lima-de-Faria, Amst., 1969.
В. Я. Бродский.

Рис. 1. Общий вид эпителиальной клетки животного при различном увеличении: а — в оптический микроскоп; б — при малом увеличении электронного микроскопа; в — при большом увеличении. Структуры ядра: 1 — ядрышко; 2 — хроматин (участки хромосом); 3 — ядерная оболочка. Структуры цитоплазмы: 4 — рибосомы; 5 — гранулярная (покрытая рибосомами) эндоплазматическая сеть; 6 — гладкоконтурная сеть; 7 — комплекс Гольджи; 8 — митохондрии; 9 — мультивезикулярные (многопузырьковые) тела; 10 — секреторные гранулы; 11 — жировые включения; 12 — плазматическая мембрана; 13 — десмосома.

Рис. 2. Схема строения клетки образовательной ткани (меристемы) растения: 1 — клеточная стенка; 2 — плазмодесмы; 3 — плазматическая мембрана; 4 — эндоплазматическая сеть; 5 — вакуоли; 6 — рибосомы; 7 — митохондрии; 8 — пластида; 9 — комплекс Гольджи; 10 — оболочка ядра; 11 — поры в ядерной оболочке; 12 — хроматин; 13 — ядрышко.

Рис. 3. Разнообразие клеток высших растений: а, б — меристематические клетки; в — крахмалоносная клетка из запасающей паренхимы; г — клетка эпидермиса; д — двуядерная клетка секреторного слоя пыльцевого гнезда; е — клетка ассимиляционной ткани листа с хлоропластами; ж — членик ситовидной трубки с клеткой-спутницей; з — каменистая клетка; и — членик сосуда.
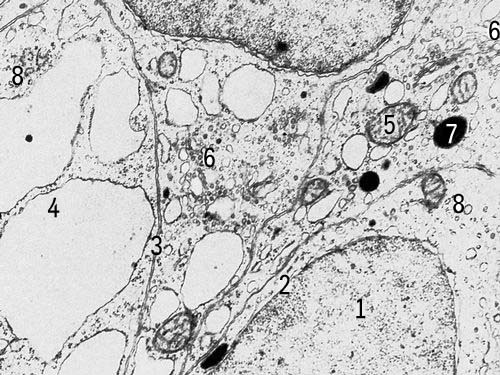
Клетки щитовидной железы крысы с включениями (увеличено в 18000 раз). Условные обозначения: 1 — ядро, 2 — ядерная оболочка, 3 — клеточная оболочка, 4 — эндоплазматическая сеть, 5 — митохондрии, 6 — комплекс Гольджи, 7 — плотные тела, 8 — рибосомы.
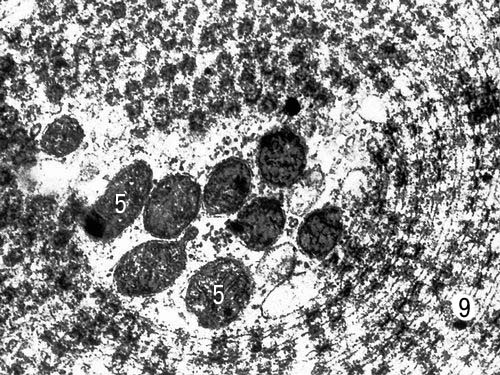
Специализированная форма мембран (пористые пластинки) в цитоплазме созревающей яйцеклетки севрюги (увеличено в 35000 раз). Условные обозначения: 5 — митохондрии, 9 — пористые пластинки.
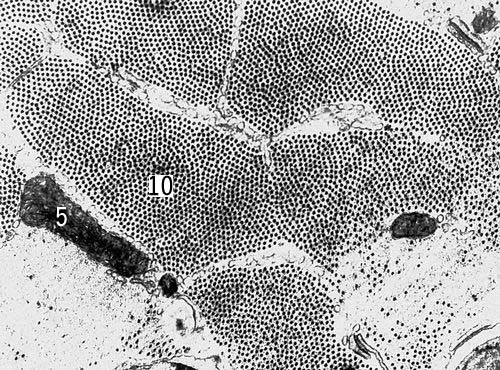
Поперечный срез мышечных клеток саранчи (увеличено в 54000 раз). Условные обозначения: 5 — митохондрии, 10 — миофибриллы.
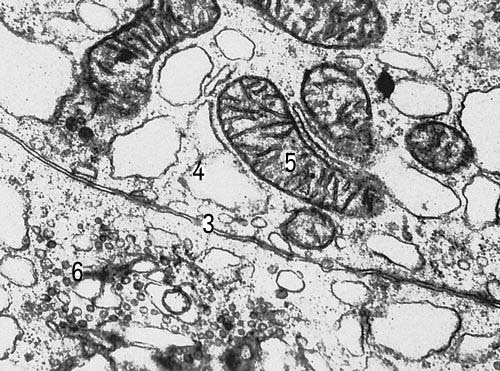
Участки двух клеток щитовидной железы крысы (увеличено в 30000 раз). Условные обозначения: 3 — клеточная оболочка, 4 — эндоплазматическая сеть, 5 — митохондрии, 6 — комплекс Гольджи.
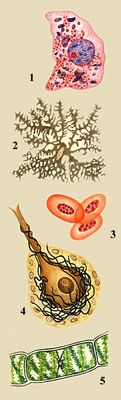
Разнообразие животных и растительных клеток: 1 — клетка печени аксолотля, в цитоплазме — красные митохондрии и фиолетовые белковые включения, в ядре — красное ядрышко и синие глыбки хроматина; 2 — хроматофор аксолотля, заполненный гранулами пигмента; 3 — эритроциты лягушки; 4 — клетка Пуркине мозжечка крысы; 5 — клетка водоросли спирогиры.
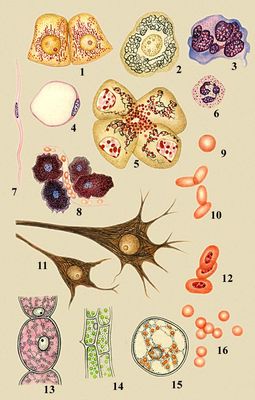
Разнообразие животных и растительных клеток: 1 — клетки почки лягушки, видны митохондрии; 2 — чувствительная клетка спинномозгового ганглия человека, виден комплекс Гольджи; 3 — мегакариоцит из костного мозга человека; 4 — жировая клетка из подкожной клетчатки крысы; 5 — клетки поджелудочной железы человека, видны комплекс Гольджи и секреторные гранулы; 6 — нейтрофильный лейкоцит человека; 7 — гладкая мышечная клетка кишечника человека; 8 — тучные клетки в рыхлой соединительной ткани крысы; 9 — эритроциты человека; 10 — эритроциты верблюда; 11 — малая и большая пирамидальные клетки коры головного мозга человека; 12 — эритроциты курицы; 13 — клетка волоска тычиночной нити традесканции; 14 — клетки листа элодеи; 15 — клетка плода ландыша; 16 — эритроциты свиньи.
Большая советская энциклопедия. — М.: Советская энциклопедия. 1969—1978.