MYRIAPODES
Les Myriapodes sont des Arthropodes terrestres antennifères (une paire d’antennes) et mandibulates possédant un grand nombre d’anneaux pédifères (plus de trois) groupés deux à deux d’une manière plus ou moins apparente (diplopodie).
Le terme fut créé par Pierre André Latreille (1796) pour la «légion» des Myriapodes, mais ce n’est qu’en 1814 que W. Leach élève les Myriapoda au rang de classe, au même niveau que les Crustacés, les Arachnides et les Insectes.
La plupart des auteurs modernes ont prétendu qu’à l’intérieur de la classe des Myriapodes il n’y avait aucune parenté étroite entre les quatre composants qui sont les ordres des Diplopodes, des Symphyles, des Pauropodes et des Chilopodes. Ils ont mis en doute la validité de ce grand groupe, considéré par eux comme non naturel, et ont élevé les quatre ordres au rang de classes, regroupées en deux séries phylétiques suivant que les gonoductes débouchent en avant du corps (progonéates) ou en arrière (opisthogonéates).
On sait aujourd’hui que la position progonéate du gonopore est secondaire, au moins chez les Symphyles: Hanseniella agilis est d’abord opisthogonéate, puis devient progonéate par développement de gonoductes d’origine ectodermique. Cette position du gonopore serait une adaptation secondaire à l’anamorphose (Tiegs); elle n’aurait donc pas de valeur phylétique.
À la lumière de travaux morphologiques récents qui ont mis en évidence un caractère commun à toute la classe des Myriapodes, à savoir la diplopodie, on peut affirmer que l’hétérogénéité de cette classe est plus apparente que réelle.
1. Caractères généraux
La diplopodie est une forme de métamérie caractérisée par la présence, plus ou moins apparente, de groupements des métamères par deux. Chacun de ces groupements, appelés diplosegments (fig. 1), porte dans le cas le plus simple, celui des Diplopodes, deux paires de pattes. Chez les Chilopodes, dont les anneaux ont une seule paire d’appendices (fig. 2), la structure bimétamérique, extérieurement inapparente, reste profonde et groupe ces anneaux par deux. La diplopodie est l’unité articulaire du corps des Myriapodes .
La séparation entre thorax et abdomen est indistincte sauf chez les Diplopodes, dont on considère comme thoraciques les quatre premiers diplosegments, apparemment simples puisque ne portant qu’une seule paire de pattes; leur métamère postérieur est considérablement réduit (pas d’appendices). Cette réduction en volume se retrouve tout le long du corps chez les Chilopodes, mais généralement sans disparition des appendices; les segments macrotergaux, plus grands, et les segments microtergaux, plus petits, alternent régulièrement. Les séquences bisegmentaires des Chilopodes avec un métamère macrotergal antérieur et un métamère microtergal postérieur sont comparables aux diplosegments des Diplopodes.
La diplopodie résulte-t-elle de l’association de simples segments par groupes de deux ou, au contraire, dérive-t-elle d’une fragmentation particulière du corps? Les travaux embryologiques de Pflugfelder sur Platyrrhacus amauros sont en faveur de la seconde explication. La segmentation du tronc se conçoit comme une fragmentation en macrosomite de premier ordre, puis en macrosomite de deuxième ordre, puis de troisième ordre (diplosegments), enfin en microsomites ou segments simples. La morphologie confirme ce processus si l’on admet que chaque macrosomite garde une autonomie dans son potentiel de fragmentation et par conséquent de réduction métamérique en volume et en nombre. Ces phénomènes aident à comprendre la réduction segmentaire localisée dans le thorax des Diplopodes (quatre diplosegments, donc quatre macrosomites de troisième ordre), celle constatée tout le long du corps des Chilopodes (métamère postérieur réduit de chaque unité bisegmentaire) et l’absence complète, ou quasi complète, du huitième métamère pédifère des Chilopodes (il n’y a pas d’appendices; réduction de l’extrémité d’un macrosomite de premier ordre qui est égale à huit microsomites). Toutes les «anomalies» de segmentation des Myriapodes peuvent ainsi être expliquées.
La notion de réduction métamérique en volume, entraînant une réduction en nombre, amène tout naturellement à évoquer ces problèmes du point de vue évolutif. Il existe deux principes évolutifs, dits de contraction (Brölemann) et d’élongation (Verhoeff), suivant que l’on considère que les Myriapodes ont perdu des segments ou bien en ont acquis au cours de leur évolution.
L’interprétation donnée ci-dessus indique qu’ils ont perdu des métamères et que les Myriapodes les plus évolués sont ceux qui possèdent le plus petit nombre de segments. Pourtant, il semble exister deux sortes de réduction métamérique qu’il est important de ne pas confondre: l’une d’ordre évolutif, fixée dans le patrimoine héréditaire, l’autre d’ordre écologique, liée à la croissance postembryonnaire et soumise aux conditions du milieu (oligomérie écologique).
2. Classification
Diplopoda
De grande taille, jusqu’à 30 cm, les Diplopoda (type: Iule) ont des téguments calcifiés et des gonopores pairs, en avant du corps (progonéates). La tête est perpendiculaire à l’axe du corps; les ocelles peuvent être absents. L’organe temporal est généralement présent; les mandibules sont divisées en partie basilaire (cardo, stipes) et partie distale (prémandibule). Les maxilles (une ou deux paires) sont soudées en gnathochilarium. Les très nombreux anneaux (11 à 80-190), sans tergites individualisés, portent deux paires de pattes s’articulant ventralement et une paire de glandes répugnatoires. Ils possèdent des organes copulateurs (gonopodes).
Chez les Pénicillates (sous-ordre, type: Polyxène), les téguments sont mous, les plaques tergales portent de grosses soies creuses (trichomes) rassemblées en touffes. Ils n’ont pas de gonopodes. Leur taille est réduite à quelques millimètres.
Symphyla
Les Symphyla sont de petite taille (quelques millimètres; type: Scutigérelle). Les téguments sont mous, les gonopores pairs en avant du corps (progonéates). La tête est perpendiculaire à l’axe du corps; il n’y a pas d’ocelles mais un organe temporal; les mandibules, à capitulum mobile, sont divisées. Il y a deux paires de maxilles: une paire libre latérale et une paire unie en labium. Ils sont ectotrophes et possèdent une paire unique de stigmates céphaliques, des filières, douze paires de pattes s’articulant ventralement pour dix-sept à vingt-quatre plaques tergales sans rapport avec des tergites segmentaires.
Pauropoda
Les Pauropoda sont de très petite taille (1 mm); ils ont des téguments mous, un gonopore impair en avant du corps (progonéates). La tête est perpendiculaire à l’axe du corps; il n’y a pas d’ocelles; une paire d’aires vitreuses latérales (pseudoculus) équivaut peut-être à l’organe temporal. Les mandibules sont simples; un hypostome ferme la capsule céphalique, avec deux paires de pointes (galea, maxilla) représentant peut-être les deux maxilles. Ils sont fortement endotrophes. Les antennes, biramées, portent un organe sensoriel globuleux original (globulus). Dix ou onze paires de pattes ambulatoires s’articulent ventralement pour six plaques tergales.
Chilopoda
Les Chilopoda, de grande taille, jusqu’à 25 cm, ont des téguments mous, un gonopore impair à l’extrémité du corps (opisthogonéates). La tête est aplatie, dans l’axe du corps, les antennes simples. Les ocelles ne sont pas toujours présents, l’organe temporal l’est généralement; les mandibules sont simples. Il existe deux paires de maxilles soudées deux à deux. La première paire de pattes est céphalisée et transformée en crochets à venin (forcipules). Les segments simples, à une paire de pattes, s’articulent latéralement. Il y a de 15 à 23 paires de pattes et jusqu’à 177 paires chez les Géophiles.
On distingue quatre sous-ordres: Géophilomorphes, Scolopendromorphes, Lithobiomorphes, Scutigéromorphes.
3. Anatomie et physiologie
Le plan d’organisation (fig. 3) est typiquement arthropodien.
Alimentation, digestion
Les Diplopodes sont surtout végétariens. On en a vu cependant ingérer des tissus animaux vivants ou frais, des cadavres de petits mammifères, des excréments. En absorbant des matières organiques en décomposition, ils peuvent jouer un rôle d’agent de dissémination de parasites par exemple. Détritivores ou saprophages, ils interviennent directement dans le processus de décomposition, le renouvellement de la matière organique et le recyclage de certains éléments minéraux dans le sol. Leur rôle est important comme fragmentateurs et transformateurs de la matière organique végétale surtout dans les premières étapes de la décomposition.
Les aliments sont déchiquetés en lanières et humectés par les sécrétions des glandes salivaires. La digestion s’opère dans l’intestin moyen qui est, en outre, la seule région absorbante du tube digestif. Les résidus sont abondants, mélangés à des particules minérales. Les déjections sont utilisées par quelques espèces dans la construction des nids de ponte et de mue.
Les Chilopodes sont des carnivores et jouent un rôle dans la régulation des populations (Collemboles par exemple). Les proies sont tuées par le venin injecté par les forcipules. Les Géophilomorphes se lovent autour de leur proie pour l’immobiliser, alors que les Scolopendromorphes utilisent à cet effet leurs pattes postérieures différenciées en pattes ravisseuses.
Une membrane péritrophique enveloppe les résidus de la digestion. C’est une sorte de gelée, dont la consistance croît d’avant en arrière chez Lithobius .
Les Pauropodes se nourrissent de substances fluides; ils ne peuvent que sucer ou piquer; ils doivent aspirer les sucs végétaux. La paroi de l’intestin moyen présente des cellules à ferments et des cellules absorbantes. L’absorption s’effectue selon des processus chimiques rappelant ceux découverts par Millot chez les Arachnides. Le liquide absorbé subit des transformations chimiques qui aboutissent à la séparation des produits assimilables et des produits de déchets; ces derniers sont rejetés dans la lumière intestinale.
Excrétion
L’excrétion s’effectue au niveau des reins ouverts (labiaux, Malpighi) et des reins fermés (athrocytes, corps adipeux, cellules péricardiales, filaments de Kowalewski chez les Chilopodes). Les embryons des Symphyles ont une glande coxale prémandibulaire, organe transitoire disparaissant après l’éclosion. Les cellules des tubes de Malpighi ont un pouvoir oxydant, ce qui est une exception dans le règne animal.
Les Myriapodes sont uricotéliques et ammoniotéliques: 20 à 40 p. 100 de l’azote total excrété pendant vingt-quatre heures est rejeté sous forme d’ammoniaque. L’origine des deux catabolites terminaux, acide urique et ammoniaque, n’est pas totalement élucidée.
Respiration, circulation
Les Myriapodes respirent généralement par des trachées. Le système trachéen est pratiquement inexistant chez les Symphyles où la respiration cutanée est très poussée; elle est même exclusive chez les larves dont les stigmates céphaliques, seuls orifices respiratoires, sont fermés. Le faible développement des trachées est, sans doute, l’une des causes de l’hygrophilie des Symphyles, puisque seul l’oxygène dissous peut traverser la paroi de l’organe respiratoire. Chez les Diplopodes et les Chilopodes, les stigmates (pairs) se situent soit sur chaque paire de pattes, soit dans les pleures. Les trachées se terminent par des trachéoles en cul-de-sac logées dans le cytoplasme des cellules trachéolaires.
Les Scutigères (Chilopodes) ont un appareil respiratoire composé d’un grand nombre de trachées libres rassemblées en deux lobes baignant dans une lacune sanguine et débouchant dans une chambre respiratoire impaire. Les passages intermittents du sang (systole-diastole) dans les espaces intertrachéens correspondent à une inspiration et une expiration. Chez les autres Myriapodes sans mouvements respiratoires, ce sont les pulsations du cœur (90 à 200 par minute chez Scutigera , 44 chez Strongylosoma ), le mouvement des muscles, etc., qui favorisent la ventilation trachéenne.
Chez les Diplopodes, la consommation d’oxygène est variable suivant les espèces, les saisons, l’activité et la nutrition, mais elle reste sensiblement égale pour les deux sexes d’une même espèce; elle diminue pendant la période d’intermue. L’oxygène consommé varie également avec le poids. Le quotient respiratoire le plus bas s’observe pendant la mue; on a attribué ce phénomène à la fixation du gaz carbonique et à son accumulation dans le tégument sous forme de carbonate de calcium.
Le système circulatoire est classique, avec un cœur dorsal, organe hématopoïétique, un péricarde, des lacunes sanguines autour de la chaîne nerveuse ventrale, des artères, etc. Chez Scutigère, il existe deux ampoules pulsatiles en avant du cœur. L’humidité, et non la nutrition, influe sur la masse sanguine et le nombre des globules, parmi lesquels il existe des éléments phagocytaires.
4. Reproduction
Biologie sexuelle
Les sexes sont séparés et seule une race de Pénicillates est parthénogénétique. Dans plusieurs espèces, le pourcentage des mâles diminue suivant les régions géographiques considérées (spanandrie).
Il existe deux types de spermatogenèse: l’un aboutissant à une seule catégorie de spermatozoïdes (Diplopodes); l’autre, selon un processus de double spermatogenèse, produisant deux spermatozoïdes, un petit et un grand (Chilopodes, Symphyles). Cette différence de taille peut être due à la présence d’un hétérochromosome dans les spermatozoïdes de grande taille; les géants donneraient des femelles, les nains des mâles. Chez les Symphyles, pourtant, la lignée d’aspect dégénératif assurerait la survie des spermatozoïdes fonctionnels dans le spermatophore.
La fécondation s’effectue dans l’oviducte, mais chez les Symphyles c’est la femelle qui féconde elle-même ses propres œufs lors de la ponte, en utilisant les spermatozoïdes stockés dans des poches de la cavité buccale. La fécondation est indirecte et externe. C’est le type le plus primitif qui soit réalisé chez les Arthropodes terrestres.
L’accouplement n’existe que chez les Diplopodes. Le mâle fabrique les spermatophores chez les autres Myriapodes.
Accouplement
En arrière de la deuxième paire de pattes ambulatoires débouchent les conduits génitaux pairs; il existe à leur orifice des organes complexes: vulves et pénis qui sont des appendices ambulatoires modifiés. Les gonopodes recueillent le sperme, issu du «pénis», grâce à une pièce spéciale, creusée en gouttière, qui le prélève et le transporte sur les vulves. Les gonopodes résultent de la transformation des huitième et neuvième paires de pattes du septième anneau, ensemble ou séparément (fig. 4). Chez les Spirostreptoidea, la gouttière est munie d’une pièce en piston, glissant dans la rainure, aidant ainsi à l’éjaculation du liquide.
Chez les Glomérides, le comportement est très particulier et même unique. Il y a accouplement en ce sens que le mâle saisit les vulves de la femelle avec ses télopodes pour la maintenir, mais il utilise, pour le transport du sperme, une boulette de fèces ou de terre modelée pendant l’accouplement. La boulette équivaut à un spermatophore sur lequel du sperme est déposé; ce dernier est ensuite introduit dans les vulves par les pièces des télopodes (Haacker).
Reproduction par spermatophore
Le spermatophore est généralement déposé, en présence de la femelle, sur une toile filée par le mâle. La succession des mouvements est une véritable danse chez Scutigère. Le mâle dirige généralement la femelle sur le spermatophore dont la fabrication s’effectue progressivement dans les voies génitales postérieures. L’appareil, complexe, comprend trois enveloppes chez les Scolopendromorphes; toutes présentent un point de moindre résistance correspondant à une invagination qui pénètre par évagination dans les voies génitales femelles; il éclate et projette le sperme sous la pression de l’enveloppe externe gonflée au contact du mucus de la région génitale.
Les Polyxènes mâles tissent une toile complexe, déposent deux gouttes de sperme au centre et laissent une double rangée de fils parallèles. Ce tissage spécial, conducteur chimique, guidera les femelles de passage qui suivront alors scrupuleusement le chemin soyeux du mâle jusqu’à la semence. Chaque mâle de passage renouvellera celle-ci. Il se produit là une entreprise collective de la part des mâles, résultant d’un certain comportement social correspondant au mode de vie en groupe. Une femelle parthénogénétique ne réagira naturellement pas au contact de la toile; elle la franchira par un côté quelconque.
Chez les Symphyles, la femelle stocke dans des réservoirs spéciaux, les poches gnathales, les spermatozoïdes, qui seront utilisés ultérieurement, d’une façon originale, pour la fécondation externe de l’œuf.
Ponte, instinct maternel
La ponte a lieu souvent dans les fentes du sol, en avril-mai dans les régions tempérées.
Les femelles de Diplopodes surveillent rarement leurs pontes (40 à 1 200-1 300 œufs suivant les espèces), mais elles les mettent au moins à l’abri soit dans un cocon de soie produite par les glandes séricigènes lorsqu’elles existent (Nématophores), soit dans des oothèques terreuses. Chez les Polydesmides, ce sont de véritables constructions en cloche avec cheminée centrale d’aération. Les parois sont crépies par un ciment fluide provenant du tube digestif. L’enveloppe sert de premier aliment aux jeunes larves. Les Iulides crépissent de même les parois d’une cavité préformée.
Les Spirobolides et les Glomérides enrobent les œufs séparément dans une capsule et les abandonnent sur le sol.
Quelques Diplopodes tissent un cocon dans lequel s’enferme la femelle. Cas unique, chez une espèce du Japon, le mâle se saisit des œufs un à un au fur et à mesure de leur ponte et surveille ensuite la couvée.
Chez les Chilopodes, la Scolopendre s’enroule autour de sa ponte pour la protéger du contact du sol, des moisissures, et la défendre contre les prédateurs. Lithobius confectionne des oothèques terreuses à l’aide des griffes en pelle des gonopodes femelles.
5. Développement et croissance
On reconnaît deux sortes de développement postembryonnaire: l’un par épimorphose, selon lequel la larve naît avec un nombre de segments pédifères fixés dès l’éclosion; l’autre par anamorphose – le plus fréquent –, au cours duquel les segments sont acquis au fur et à mesure du déroulement des stades de croissance jusqu’à l’état adulte. Dans les deux cas, le nombre définitif des segments est rarement fixé chez l’adulte; il est variable, dans de larges proportions (type anomoméristique), chez les individus d’une même espèce.
La première larve des Diplopodes, Symphyles et Pauropodes est hexapode, et chez la plupart des groupes le premier stade est précédé d’un stade pupoïde, où la larve est enveloppée dans une membrane embryonnaire sans trace de segmentation mais parfois avec des soies et l’organe d’éclosion (dent ou épine).
Le nombre des stades larvaires n’est pas uniforme; il varie chez les Diplopodes suivant que le nombre des anneaux est fixe ou variable chez l’adulte et suivant que le nombre de ces anneaux est faible ou élevé. Le rythme de croissance est irrégulier; rapide au début, il ralentit vers la fin, mais les larves d’une espèce donnée de Iulides n’ont pas toutes le même rythme; elles sont soit en avance, soit en retard par rapport au rythme moyen, ou les unes par rapport aux autres.
Chez Glomeris (Diplopode), à la période anamorphe, créatrice de segments, succède une période épimorphe au cours de laquelle sont acquis des caractères autres que la segmentation. On est ainsi en présence d’un cas strictement comparable à celui des Chilopodes, Lithobiomorphes en particulier, éminemment évolués. Une contraction tachygénétique du développement postembryonnaire tend à réduire le nombre des stades, entraînant une diminution du nombre des anneaux définitivement acquis par l’adulte.
L’épimorphose se rencontre chez les formes primitives des Chilopodes à grand nombre de segments (Géophilomorphes, Scolopendromorphes). La croissance embryonnaire est prolongée par rapport aux anamorphes, qui naissent avec un nombre plus faible de segments. Le premier stade des Géophilomorphes est dit péripatoïde; le tube digestif contient du vitellus et la segmentation n’est pas achevée. Chez les Scolopendromorphes, le stade péripatoïde a disparu, le développement est plus avancé.
L’anamorphose est le fait des Lithobiomorphes et des Scutigéromorphes, chez lesquels la première larve correspond à un véritable embryon par comparaison aux Géophilomorphes; elle n’a que sept paires de pattes au lieu de quinze chez l’animal complet.
Le cycle biologique se déroule sur un ou deux ans (Polydesmides, Craspédosomides). Cependant, il est beaucoup plus lent (plusieurs années) chez les autres Diplopodes (Iulides). Les populations présentent alors des structures démographiques instantanées complexes car, à un instant donné, elles sont constituées d’un ensemble d’individus provenant de générations différentes.
Zone de croissance
Les nouveaux segments s’élaborent progressivement dans une zone interne comprise entre le dernier anneau pédifère et le telson. Chez les Diplopodes, l’anneau franchit quatre étapes. D’abord interne, d’embryosomite l’anneau devient un éosomite comprimé. Exprimé extérieurement, il est apode (mésosomite) mais contient, sous forme de bourgeons, les futures pattes de l’anneau complet. Tous les éléments sont libérés en bloc à chaque mue. Lorsque la croissance s’achève, tous les éléments sont encore présents, mais ils ne verront jamais le jour, en principe. Pourtant l’activité de la zone de croissance peut reprendre et de nouveaux segments peuvent apparaître dans certaines conditions. Cela est lié à un phénomène complexe, encore mal défini, propre à quelques espèces d’Iulides, la périodomorphose, découverte par Verhoeff. L’adulte mâle, sa fonction de procréateur remplie, mue et devient un «mâle intercalaire» dont les gonopodes sont réduits et ressemblent à ceux des larves immatures; puis il mue pour donner naissance soit à un nouveau mâle intercalaire, soit à un mâle apte à copuler.
La périodomorphose pourrait n’être qu’un phénomène biologique particulier lié à des activités hormonales cycliques, où intervient, notamment, une hormone androgène commandant les caractères sexuels secondaires (évolution des caractères gonopodiaux) en dehors de toute considération de croissance segmentaire. On peut envisager qu’une activité cyclique parallèle et concomitante de la gonade se produit avec des périodes de repos spermatogénétique (adulte) et d’activité chez les intercalaires préparant le futur stade «adulte».
La périodomorphose, pour Brölemann, est le reliquat d’un mode de croissance ancestral, une sorte de résurgence qui se produit dans certaines conditions.
Le déterminisme de la mue est sous l’influence d’un système (cellules neurosécrétrices-glandes endocrines) qui n’est pas sans analogie avec celui des Insectes. Un complexe est inhibiteur, un second stimulateur.
6. Éthologie et écologie
Réactions aux stimuli
Tact
La sensibilité tactile est d’autant plus grande que l’animal est plus agile; les soies des antennes, organes récepteurs, sont d’autant plus abondantes que le sujet est plus actif. Elles fournissent des renseignements pour l’orientation: sans antennes, l’animal ne peut franchir un obstacle.
Chez les Pauropodes, le tact dépend d’organes spéciaux, les trichobothries, longs poils mobiles au fond d’une dépression tégumentaire caliciforme.
À une pression sur le corps, les Diplopodes répondent par un enroulement ou une immobilisation réflexe semblable à la simulation de la mort de nombreux Arthropodes.
Les Chilopodes ont un stéréotropisme accentué (besoin d’un contact avec un corps dur).
Odorat, goût
Le siège de l’olfaction serait dans les chevilles des antennes et les quilles sensorielles de l’extrémité. Pour certains auteurs, ce serait également l’organe temporal, par analogie avec des organes sensoriels des Insectes dont le rôle olfactif a été prouvé. L’animal est surtout guidé par l’odorat pendant la marche; il explore le sol avec les quilles du dernier article antennaire.
L’odorat des Chilopodes est très développé; cependant, Lithobius n’est guidé vers sa nourriture ni par la vue ni par l’odorat, mais plutôt par le tact: privé d’antennes, il ne peut rechercher ses aliments et meurt de faim.
Le goût a son siège à la base des quilles des palpes des mâchoires et du gnathochilarium.
Ouïe
Les organes particuliers de la hampe antennaire de Scutigère sont auditifs (Verhoeff), mais les autres Chilopodes ne semblent pas avoir de sensations auditives. Le globulus des Pauropodes est peut-être un organe auditif, son ampoule jouant le rôle de caisse de résonance.
Vue
La vision est assurée par des ocelles. Chez les Scutigères, les ocelles (100 à 600) sont groupés et l’ensemble a l’aspect d’un œil composé d’Insecte. L’ocelle ressemble à l’ommatidie des Insectes et des Crustacés (présence d’un rhabdome, d’un corps cristallin).
Les formes aveugles répondent plus rapidement que les oculées aux excitations lumineuses; les ocelles ne sont donc pas les seuls organes photorécepteurs. Les sensations sont alors dermatoptiques.
L’évagination digitiforme du septième article antennaire du Diplopode aveugle Polydesmus pourrait être l’organe photorécepteur.
La vision doit être très fruste, même chez les oculés; ils voient à courte distance et semblent mal interpréter les formes.
Autres formes de sensibilité
La réponse aux vibrations est la mise en marche, l’immobilité ou l’enroulement suivant l’intensité de l’excitation. Le siège est attribué aux poils tactiles des antennes.
La géotaxie est positive chez les Diplopodes. Les organes récepteurs pourraient être les organes temporaux qui existent chez de nombreux Diplopodes et quelques Chilopodes. Ce sont des invaginations tégumentaires de formes variables. Leur structure rappelle cependant celle d’organes olfactifs des Insectes.
La sensibilité thermique est bien développée. La vitesse de déplacement et l’activité sexuelle augmentent avec la température. On n’a pas reconnu de sensilles spécifiquement thermorécepteurs.
Attaque et défense
Les Chilopodes possèdent dans leurs forcipules un puissant instrument inoculateur de venin. Les accidents mortels chez l’Homme dus à la morsure des Scolopendres sont rarissimes et ne peuvent survenir que chez de très jeunes enfants mordus en des régions très précises du corps (tête, cou, par exemple). Les effets des morsures ont été très exagérés. Dans les régions tempérées, on distingue deux sortes de morsures: une d’hiver insignifiante, une de printemps plus sérieuse. La douleur et l’enflure peuvent être vives et importantes. Par contre, la morsure est mortelle pour les Arthropodes et quelques petits Mammifères. La Scolopendre est immunisée contre son propre venin. La quantité de venin émise est quatre fois plus petite que celle du Scorpion; il hémolyse le sang des Mammifères, et ces propriétés peuvent être exaltées par l’adjonction de substances lipoïdes.
Les Géophilomorphes sécrètent en outre, par leurs glandes ventrales, un liquide gluant renfermant de l’acide cyanhydrique; les petits Arthropodes y sont sensibles.
Les Diplopodes, longtemps considérés comme inoffensifs, peuvent être dangereux aussi bien pour l’Homme que pour les animaux en raison des poisons qui s’écoulent par les pores (une paire par anneau) des glandes répugnatoires. Les différents principes actifs du venin semblent être spécifiques des groupes systématiques. Ce sont l’acide cyanhydrique associé à des produits dérivés d’un glucoïde par hydrolyse chez les Polydesmides, des quinones irritant la peau chez les Iuloïdes, de la glomérine et de l’homoglomérine chez les Glomérides.
La substance sécrétée par les Polydesmoïdes est naturellement toxique et les indigènes du Mexique en utilisent des extraits, associés à ceux de plantes, pour empoisonner leurs flèches. Un petit Polydesmide du Soudan est aussi riche en HCN que les plantes cyanogènes les mieux pourvues (de 57,3 mg à 89,77 mg pour 100 individus). Les organes de décharges cyanogénétiques comportent un réservoir spécial, ou vestibule, dans lequel la sécrétion de la glande, composé cyanogène inoffensif, est décomposée par un facteur chimique au moment de la décharge. Cela explique pourquoi les Polydesmes, sensibles au HCN, sont capables d’émettre une substance toxique sans en être incommodés.
Certains Iulides, pourtant non cyanogènes, sont dangereux pour l’Homme par leur capacité de projeter un liquide corrosif à 60-80 cm de distance. On connaît plusieurs cas de cécité momentanée ou définitive par projection de benzoquinones dans les yeux.
La fuite est bien souvent le seul moyen d’échapper à l’ennemi, mais le déplacement est d’autant plus rapide que le nombre de pattes est moins élevé. La locomotion des Myriapodes est bien particulière surtout chez les Diplopodes dont les pattes sont très nombreuses. Les mouvements successifs déterminent des vagues métachronales qui progressent le long du corps. La coordination des mouvements est contrôlée par un mécanisme nerveux selon lequel l’excitation voyage le long de la chaîne nerveuse et engendre une série de mouvements produits par les impulsions des neurones moteurs des ganglions successifs.
La vague métachronale des Chilopodes est moins perceptible à l’œil (nombre de pattes plus faible), mais le corps ondule par une différence de phases existant entre les pattes propulsives d’un côté et de l’autre du corps (fig. 6).
Du point de vue de l’orientation et du maintien d’une course déterminée, les excitations de provenance extérieure sont intégrées dans des centres nerveux suivant un système à afférence résiduelle ou réafférence. Un programme établi dans un centre supérieur commande à des centres inférieurs où les données afférentes et efférentes sont traitées. Ces données (excitations plus ou moins fortes) totalisées sont mises en balance et leurs «résidus» transmis au centre supérieur directeur.
Certains Pénicillates (Synxenoïdes) peuvent sauter à l’aide de leurs dernières paires de pattes modifiées.
Pour se défendre, certains Myriapodes, comme les Diplopodes Gloméroïdes, émettent des sons. Peut-être s’agit-il encore d’une sorte de comportement sexuel car on a montré récemment que la stridulation chez certains Gloméroïdes est un prélude à l’accouplement. Un Arthrosphæra indien vibre nettement lorsqu’on le saisit. Une espèce de Madagascar porte le nom évocateur de musicus .
L’émission des sons chez les Chilopodes est moins évidente. Ce sont les mouvements spasmodiques des pattes postérieures autotomisées de Rhysida nuda qui produisent des sons. Le son criard produit par les membres détachés d’une grande Scutigère de Malaisie est particulièrement efficace puisqu’il fixe l’attention des prédateurs.
L’autotomie, accompagnée de sons ou non, est un réflexe que l’on déclenche facilement par des excitations de toutes sortes. Elle est d’autant plus développée que l’animal est agile. Ainsi peut-on obtenir la perte totale des appendices chez la Scutigère qui est d’une agilité extrême.
Émission de lumière
Quelques Chilopodes, et plus particulièrement les Géophilomorphes, laissent sur leur passage des traces lumineuses bleuâtres ou verdâtres suffisamment intenses, dit-on, pour permettre de lire le journal à leur clarté.
On n’a pas pu déceler la présence de véritables organes lumineux ou de bactéries lumineuses. L’émission de lumière est sans doute due à des phénomènes d’oxydation. Un Diplopode des États-Unis, Luminodesmus , luit dans l’obscurité sur toute la surface de son corps. On n’a pas encore expliqué ce comportement particulier; on peut seulement dire qu’il ne s’agit pas de reconnaissance des sexes.
Activité constructrice
Pour muer ou pour protéger leurs pontes, de nombreux Myriapodes élaborent des constructions variées qui tendent à montrer un perfectionnement graduel de l’activité constructrice. On peut reconnaître plusieurs degrés de perfectionnement, allant de l’aménagement de la cavité préformée jusqu’à l’élaboration d’un édifice plus complexe, faisant appel à des capacités psychiques de plus en plus grandes. La logette d’un Polydesmide exotique, par exemple, possède un clapet interne destiné à régulariser le degré hygrométrique de la chambre et à faire obstacle à l’intrusion des prédateurs.
Habitat
Les Myriapodes sont hygrophiles et lucifuges; on les rencontre en particulier dans les fentes du sol, les litières forestières, sous les pierres, les bois pourris. Leur hygrophilie est variable d’une espèce à l’autre; d’une manière générale, les Diplopodes sont plus hygrophiles que les Chilopodes; quelques-uns se trouvent dans les endroits sablonneux, les buissons, dans les arbres, tandis que d’autres pénètrent spontanément dans l’eau.
L’abondance des Myriapodes au niveau du sol varie suivant sa nature (sable, argile, calcaire), les saisons, mais aussi suivant les espèces.
La température joue un grand rôle dans la distribution; les Chilopodes sont eurythermes et quelques espèces se rencontrent un peu partout, aussi bien en plaine qu’en montagne. Il y a peu de formes adaptées à l’altitude et le nombre des espèces diminue au fur et à mesure que l’on s’élève.
Quelques Myriapodes ont colonisé certains milieux particuliers. Les nidicoles sont des formes banales de surface qui ont colonisé ces microcavernes que représentent les nids d’oiseaux, de petits mammifères. Les Myriapodes zoophiles peuvent être considérés comme des inquilins, mais il existe quelques espèces de Diplopodes qui sont de véritables myrmécophiles ou termitophiles.
Les cavernicoles vrais sont spécialement adaptés aux conditions particulières de vie dans les galeries profondes et obscures. D’autres espèces ne sont pas inféodées à ce milieu où elles pénètrent pourtant.
La zone subterrestre des littoraux marins a été colonisée par des Chilopodes thalassophiles, Géophilomorphes en particulier, qui vivent dans les graviers, les fentes des falaises, les trottoirs à Lithothamnium , les débris végétaux et se laissent recouvrir par la marée.
Pseudoparasitisme
Dans le cas des fosses nasales et des oreilles de l’Homme, ce sont généralement des Chilopodes Géophilomorphes, dont le corps est filiforme, qui s’introduisent dans les sinus pendant le sommeil. Il s’agit d’un cas de pseudoparasitisme facultatif, qui est peut-être un essai d’adaptation à la vie parasitaire dans des conditions précises (parasitisme inchoatif de Giard); il ne faut pas le confondre avec la présence de Chilopodes ou de Diplopodes dans le tube digestif humain; ils sont avalés, dans ce cas, avec des aliments.
Rôle économique
Les Diplopodes, et plus encore les Symphyles, ont un rôle économique important par leur alimentation végétale. Ils s’attaquent aux plantes cultivées. Ce sont de véritables fléaux dans certaines régions et pour certaines cultures (betterave, maïs).
Néanmoins, ils ne sont pas toujours responsables des méfaits qui leur sont imputés car ils s’introduisent dans un cycle où les premiers agents pathogènes du sol, les parasites réels, etc., occasionnent une nécrose des tissus. C’est à partir de ces tissus nécrosés que Blaniulus guttulatus agit, par exemple. La présence de «ce grand destructeur de pommes de terre ou de fraises» se constate après une première infestation. Les cultures intensives, leur mécanisation et l’emploi exagéré des pesticides et de désherbants, qui perturbent l’équilibre biologique du sol, jouent également un grand rôle dans le développement des attaques.
Comme les vers de terre, les Myriapodes contribuent à un brassage du sol, hâtent la décomposition des tissus et font entrer plus rapidement dans le circuit utile les éléments que contiennent ces tissus. En outre, un gros Diplopode de Porto-Rico peut dévorer deux mille cochenilles nuisibles en trois heures.
On a signalé en Afrique, en France, en Italie et récemment au Japon, des masses impressionnantes de Diplopodes qui se déplacent. Quelquefois, ces milliers d’individus, et même ces dizaines de milliers d’individus, troublent le trafic ferroviaire, à telle enseigne que des bombardements au gas-oil enflammé peuvent être nécessaires pour dégager la voie. Au Zaïre, Zantekius weberi formait une colonne de 150 m de long sur 0,70 m de large (5 000 individus environ). Au Japon, les chiffres records de 4 000 à 10 000 au mètre carré, soit 120 à 400 millions d’individus, sont signalés. On a publié le chiffre de 20 000 millions d’individus représentant le «réservoir vivant» de la région intéressée; chiffre très supérieur aux 65 340 000 des États-Unis. En déplacement, les Diplopodes pénètrent carrément dans les rivières. On ne connaît pas encore les causes exactes de ces déplacements, mais ils coïncident avec la période de copulation et avec la saison des pluies sous les tropiques.
myriapodes [ mirjapɔd ] n. m. pl.
• 1806; de myria- et -pode
♦ Zool. Classe d'arthropodes terrestres dont le corps est formé d'anneaux portant chacun une ou deux paires de pattes. ⇒ mille-pattes. Principaux myriapodes : géophile, gloméris, scolopendre. Au sing. L'iule est un myriapode.
myriapodes
n. m. pl. ZOOL Classe d'arthropodes terrestres dont le corps est formé d'un grand nombre de segments presque identiques portant chacun une ou deux paires de pattes. (La morsure de certaines espèces de grande taille est venimeuse.)
— Sing. Un myriapode. Syn. cour. mille-pattes.
⇒MYRIAPODES, subst. masc. plur.
ENTOMOL. Classe d'animaux arthropodes terrestres dont le corps allongé est formé de nombreux anneaux portant chacun une ou deux paires de pattes. Synon. mille-pattes. Larves de myriapodes (CUÉNOT, J. ROSTAND, Introd. génét., 1936, p.67). Pareils aux scorpions blancs des cavernes, aux scorpions sans yeux, à poils tactiles, parmi les myriapodes qui mangent les champignons et les excréments des chauves-souris (ARNOUX, Paris, 1939, p.237).
Prononc. et Orth.:[ ]. Att. ds Ac. dep. 1835. LITTRÉ: ,,voyelle myriopode. Myriapode est mal fait``. Étymol. et Hist. 1806 (DUMÉRIL Zool., p.292). Empr. au lat. sc. myriapoda «id.» 1796 (LATREILLE, Précis des Caractères génériques des Insectes d'apr. AGASSIZ Myr., p.2), 1802 (ID., Hist. nat. gén. et partic. des crustacés et des insectes, t.3, p.43), formé à l'aide du gr.
]. Att. ds Ac. dep. 1835. LITTRÉ: ,,voyelle myriopode. Myriapode est mal fait``. Étymol. et Hist. 1806 (DUMÉRIL Zool., p.292). Empr. au lat. sc. myriapoda «id.» 1796 (LATREILLE, Précis des Caractères génériques des Insectes d'apr. AGASSIZ Myr., p.2), 1802 (ID., Hist. nat. gén. et partic. des crustacés et des insectes, t.3, p.43), formé à l'aide du gr.  , v. myriade et de
, v. myriade et de  ,
,  «pied»; cf. le gr.
«pied»; cf. le gr. 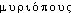 , -
, - «qui a dix mille pieds, aux pieds innombrables».
«qui a dix mille pieds, aux pieds innombrables».
]. Att. ds Ac. dep. 1835. LITTRÉ: ,,voyelle myriopode. Myriapode est mal fait``. Étymol. et Hist. 1806 (DUMÉRIL Zool., p.292). Empr. au lat. sc. myriapoda «id.» 1796 (LATREILLE, Précis des Caractères génériques des Insectes d'apr. AGASSIZ Myr., p.2), 1802 (ID., Hist. nat. gén. et partic. des crustacés et des insectes, t.3, p.43), formé à l'aide du gr. , v. myriade et de , «pied»; cf. le gr. , - «qui a dix mille pieds, aux pieds innombrables».
myriapodes [miʀjapɔd] n. m. pl.
ÉTYM. 1807; de myria-, et -pode.
❖
♦ Zool. Classe d'animaux arthropodes dont le corps est formé d'anneaux portant chacun une (chilopode) ou deux (chilognathes ou diplopodes) paires de pattes. ⇒ Mille-pattes. || Principaux myriapodes : géophile, gloméris, scolopendre. — Sing. || Un myriapode.
Encyclopédie Universelle. 2012.