ÉCOLOGIE
Le terme d’écologie (du grec oikos , demeure, et logos , science) a été proposé par E. Haeckel en 1866 pour désigner la science qui étudie les rapports entre les organismes et le milieu où ils vivent. Cette définition reste encore valable, mais elle demande à être approfondie, car l’écologie a considérablement évolué depuis ses débuts. Pour la situer par rapport aux autres sciences biologiques, il est commode de considérer les divers niveaux d’organisation de la matière vivante.
Le système le plus simple ayant toutes les caractéristiques fondamentales des êtres vivants est la cellule . Des cellules, associées en tissus et en organes, sont intégrées en organismes pluricellulaires , animaux ou végétaux.
À un niveau d’intégration supérieur, les individus de certaines espèces peuvent constituer des colonies , comme chez les Cœlentérés, ou des sociétés , comme chez les termites, les fourmis et les abeilles. À un niveau encore plus élevé se place, pour tous les êtres vivants, la population , ensemble des individus d’une même espèce liés entre eux par des liens parentaux, c’est-à-dire génétiques; selon que la reproduction est sexuée ou non, ces liens sont évidemment différents.
Les populations des diverses espèces, bactériennes, végétales et animales, groupées en un lieu déterminé, forment un niveau d’organisation plus complexe encore, la communauté biologique ou biocénose ; le milieu physique et chimique où celle-ci est installée est souvent appelé biotope et leur ensemble est un écosystème . Le niveau ultime d’organisation du monde vivant est constitué par l’ensemble des écosystèmes de la planète, c’est-à-dire l’écosphère .
La structure et le fonctionnement des cellules constituent l’objet de la cytologie et de la physiologie cellulaire; la structure et le fonctionnement des êtres pluricellulaires concernent l’histologie, l’anatomie et la physiologie des organismes. L’écologie a donc pour objet essentiel l’étude des niveaux supérieurs d’organisation de la matière vivante, de la population monospécifique à l’écosystème et à la biosphère: comme pour les niveaux inférieurs, mais cette fois à l’échelle de systèmes particulièrement complexes, il s’agit de décrire des structures, de comprendre des fonctionnements et de reconstituer des évolutions. On ne peut toutefois saisir les phénomènes jouant à un certain niveau d’intégration sans connaître ceux qui interviennent aux niveaux inférieurs: de même que l’organisation des écosystèmes ne peut être comprise qu’en tenant compte du fonctionnement des populations, celui-ci ne peut l’être sans référence aux relations que chaque individu entretient avec le milieu.
Ces relations entre les individus et le milieu où ils vivent constituent le domaine de l’auto-écologie . Elles concernent à la fois les individus, pour eux-mêmes, et la population qu’ils forment: l’auto-écologie aborde donc aussi bien des problèmes physiologiques – on parle alors d’écophysiologie – que démographiques. Dans cette perspective, le milieu est conçu comme un ensemble de facteurs dont on distingue deux types: les facteurs abiotiques , liés au milieu physique et chimique, et les facteurs biotiques , liés aux êtres vivants présents dans l’écosystème étudié.
La synécologie (parfois appelée biocénotique ) envisage essentiellement la structure et le fonctionnement des écosystèmes, mais son champ s’étend également aux complexes d’écosystèmes, associés par exemple dans un même bassin versant, et finalement à l’écosphère entière.
Discipline biologique, l’écologie ne se contente pas de décrire des structures et d’analyser des fonctionnements: elle tente de les interpréter dans une optique évolutionniste et doit considérer les systèmes qu’elle étudie – populations, écosystèmes – à différentes échelles de temps.
L’écologie est ainsi une véritable «biologie de la nature», à la fois analytique et synthétique. Son originalité, par rapport aux autres sciences de la vie, tient au niveau d’intégration élevé de ses objets d’étude, ce qui lui impose des méthodes particulières. Il existe cependant des liens très étroits entre l’écologie et, d’une part, la physiologie des organismes, d’autre part, les disciplines abordant les problèmes d’évolution (génétique des populations et biogéographie notamment).
Si l’étude des relations de l’homme avec son environnement déborde le cadre de l’écologie ainsi définie, celle-ci n’en est pas moins indispensable à la compréhension de ces relations: la connaissance du fonctionnement des systèmes écologiques et des mécanismes assurant leur stabilité fournit en effet les fondements d’une gestion rationnelle et intégrée des écosystèmes.
1. L’individu dans son environnement
Écophysiologie
Cette branche de l’écologie a pour objet d’analyser le fonctionnement de l’organisme individuel dans le cadre des contraintes que lui impose son milieu, afin de comprendre son adaptation à ces contraintes et de déterminer sa capacité à survivre lorsqu’elles changent. Du point de vue méthodologique, l’écophysiologie s’est développée selon deux directions complémentaires.
Des techniques maintenant très fines permettent des mesures de terrain précises, tant des caractéristiques de l’environnement immédiat de l’organisme (le microclimat , par exemple) que de divers paramètres indicateurs de son état physiologique. L’automatisation autorise des mesures périodiques ou continues grâce auxquelles une véritable approche du fonctionnement est rendue possible. La radiotélémétrie , du fait de la miniaturisation des matériels, offre de nombreuses possibilités dans l’étude de la physiologie in situ des organismes.
Grâce à des installations permettant la reconstitution contrôlée de nombreux facteurs abiotiques (enceintes climatiques, phytotrons), les organismes peuvent être étudiés d’une façon qui apporte aux résultats obtenus sur le terrain une sécurité complémentaire en même temps qu’elle en augmente la précision.
Cependant, certains problèmes ne peuvent guère être étudiés qu’au laboratoire et il se pose alors la question de l’extrapolation, car la transposition à la nature des résultats obtenus n’est possible qu’avec une bonne connaissance de l’environnement naturel des organismes et avec les moyens de le reconstituer de façon satisfaisante. Cela est souvent réalisable pour les facteurs abiotiques majeurs (photopériode, température et thermopériode, humidité relative, par exemple), mais il est généralement impossible de recréer l’environnement biotique, ne serait-ce qu’à cause de la complexité des régimes alimentaires naturels.
Pour l’essentiel, les recherches écophysiologiques peuvent se grouper selon deux axes principaux: l’établissement des bilans énergétiques individuels et l’étude de l’adaptation au milieu physico-chimique.
Énergétique individuelle
Tout individu, bactérien, végétal ou animal, doit incorporer une certaine quantité d’énergie pour assurer ses fonctions vitales.
Les végétaux chlorophylliens utilisent l’énergie du rayonnement solaire. Toute la lumière (LT), parvenant au contact de la plante n’est pas employée pour la photosynthèse (fig. 1 a): une partie importante n’est pas utilisée (LNU), car perdue par réflexion. La lumière effectivement absorbée (LU) n’est elle-même pas intégralement employée par la plante car une certaine fraction (LNA) est dissipée sous forme de chaleur; le reste se retrouve sous forme d’énergie chimique dans des molécules organiques (PB, photosynthèse brute). Celles-ci serviront à la synthèse des tissus et des éléments reproducteurs, où se trouve constitué ainsi un certain stock d’énergie (PN, production nette). Cependant l’élaboration de ces constituants biologiques se traduit par une dépense, qui s’ajoute à celles qu’impliquent les processus d’entretien des tissus vivants. Ces dépenses (R) sont traduites par la respiration (consommation d’oxygène et rejet de gaz carbonique). La masse de tissus nouveaux qu’un végétal élabore ainsi au cours d’un temps déterminé constitue sa production ; la production végétale est appelée production primaire .
Les facteurs exerçant une influence sur l’assimilation de l’énergie lumineuse par les plantes vertes sont nombreux. La latitude et l’exposition, par exemple, déterminent la quantité maximale d’énergie solaire susceptible d’être reçue annuellement par unité de surface, tandis que la nébulosité joue un rôle important en filtrant une partie de la lumière. La teneur de l’air en gaz carbonique, lequel est la source du carbone qu’utilise la plante, influe sur le rendement de la photosynthèse. Jouent également un rôle les particularités morphologiques des organes chlorophylliens et leur durée de vie, qui peut être de quelques mois comme chez de nombreux arbres feuillus de région tempérée, ou atteindre plusieurs années, telles les feuilles du chêne vert ou les aiguilles de nombreux résineux.
Chez les animaux (fig. 1 b), l’énergie chimique ingérée sous forme d’aliments n’est que partiellement assimilée (A); une partie (NA) est rejetée dans les excréments et l’urine (excrétats). De l’énergie assimilée A, une première partie P est stockée dans les cellules et les tissus que l’animal construit au cours de sa croissance (Pc ) et pour assurer sa reproduction (Pr ), ainsi que dans les substances que certaines espèces élaborent, comme la soie ou le mucus (Ps ): toute cette énergie est appelée la production de l’organisme (P). La seconde partie R représente l’énergie consommée au cours des synthèses organiques et, d’une façon générale, celle qu’utilise l’individu pour couvrir ses dépenses de maintenance (entretien cellulaire, activités motrices); ces dépenses sont associées aux échanges respiratoires (R).
La répartition de l’énergie ingérée entre les diverses voies ainsi définies constitue le bilan énergétique d’un individu , bilan qui peut être caractérisé par divers rendements:
– on appelle rendement écologique de croissance le rapport Pc /I, fraction de l’énergie ingérée effectivement stockée dans les tissus nouveaux; la même expression est employée pour le rapport P/I, c’est-à-dire pour la fraction d’énergie allouée tant à des tissus nouveaux qu’à la reproduction et même aux diverses sécrétions;
– le rendement de croissance tissulaire Pc /A précise la fraction des éléments assimilés qui est transformée en matière vivante et, là aussi, la même expression est souvent employée pour le rapport P/A; peut-être ce dernier mériterait-il plutôt le nom de rendement de synthèse tissulaire;
– le rendement d’assimilation A/I caractérise l’aptitude de l’individu à assimiler l’énergie chimique contenue dans ses aliments: il est généralement élevé chez les carnivores (de l’ordre de 0,80), très variable chez les herbivores (de 0,75 à moins de 0,15), faible ou très faible chez les détritivores et les géophages (de 0,10 à 0,05).
Les rendements Pc /I et Pc /A varient évidemment beaucoup avec l’âge des individus, s’annulant chez les animaux à croissance finie comme de nombreux Arthropodes, les Mammifères, les Oiseaux. Même chez les organisnes à croissance continue comme les Poissons, ces rendements diminuent sensiblement avec l’âge. D’une manière générale, ils sont bien meilleurs chez les organismes hétérothermes que chez les homéothermes qui consacrent une part importante de l’energie assimilée au maintien de la température corporelle.
L’énergie assimilée A ne peut pas être mesurée directement. Lorsque la production est négligeable, elle peut être mesurée par la chaleur dégagée par l’individu ou par son intensité respiratoire. C’est une quantité qui varie considérablement selon l’espèce, selon le sexe, l’âge et la taille des individus, selon aussi les conditions du milieu. Chez tous les animaux (fig. 2), l’énergie ainsi dépensée par unité de temps est liée au poids individuel W par une relation exponentielle du type A = K 憐 Wn , où K est une constante dépendant de l’état homéo- ou hétérotherme de l’espace et n une constante sensiblement égale à 2/3.
Les individus les plus grands ont donc, par unité de poids, un métabolisme plus faible que ceux de petite taille: leur surface corporelle proportionnellement moins importante se traduit par des déperditions d’énergie relativement moindres. Un éléphant dépense ainsi 54 kJ/g/jour, alors que chez une musaraigne cette dépense est de 3 474 kJ/g/jour. Les homéothermes ont un métabolisme bien plus actif que les hétérothermes, et qui varie de façon différente avec la température extérieure: chez les homéothermes, il présente un minimum correspondant à une température dite de neutralité thermique tandis que, chez les hétérothermes, il diminue au fur et à mesure que la température s’abaisse. Dans le cas des animaux aquatiques, le métabolisme s’accroît avec la teneur en oxygène de l’eau. Chez tous les organismes, enfin, les dépenses augmentent avec l’activité musculaire.
Adaptation au milieu physico-chimique
Notion de valence écologique
La valence écologique d’une espèce est la capacité que possède celle-ci de peupler des milieux différents. Une espèce à valence écologique élevée pourra peupler un grand nombre de milieux; elle est appelée euryèce. Une espèce à basse valence écologique, cantonnée dans un petit nombre de milieux, est dite au contraire sténoèce. Cette notion peut être considérée par rapport à tel ou tel facteur physique ou chimique: on parle ainsi d’espèce eury- et sténothermes (plus ou moins tolérantes vis-à-vis de la température ambiante), eury- et sténohygriques (vis-à-vis de l’humidité relative de l’air), eury- et sténohalines (vis-à-vis de la salinité du milieu).
En règle générale, les animaux homéothermes sont plus euryèces que les hétérothermes, car ils contrôlent mieux leur milieu intérieur et sont par suite plus indépendants du milieu environnant. Les exceptions ne manquent cependant pas; c’est ainsi que beaucoup de Mammifères de petite taille habitant le sol sont très sténoèces et ne peuvent supporter la saison froide qu’en état d’hibernation. De même les Oiseaux et les Mammifères polaires sont pour la plupart sténoèces, et plus précisément sténothermes, car ils sont incapables de résister à une température élevée. Plusieurs homéothermes équatoriaux sont également sténoèces, car ils sont liés à un intervalle limité de variations de la température et de l’humidité de l’air. En milieu marin, les espèces de haute mer sont plus sténoèces que les espèces du littoral soumises aux variations du milieu provoquées par le voisinage du continent, la faible profondeur et les marées.
Notion de facteur limitant
Pour une certaine espèce vivant dans un certain milieu, le facteur limitant est celui qui exerce l’action la plus efficace sur la présence de l’espèce: s’il tombe au-dessous d’un certain seuil (valeur minimale) ou dépasse au contraire un seuil maximal, la vie de l’espèce n’est plus possible (fig. 3). En général, plutôt que d’un seul facteur, il s’agit d’un complexe de facteurs, dominé par l’un d’eux. Ainsi, dans les déserts tropicaux comme le Sahara, l’humidité du milieu (humidité de l’air, eau disponible pour les organismes) est le facteur limitant le plus évident; cependant des relations nombreuses avec l’irradiation, la ventilation et la température peuvent en modifier le rôle et la valeur, soit dans le temps (conditions nocturnes relativement supportables), soit dans l’espace (micro-orientations favorables, protection contre les vents desséchants, etc.). Cela montre combien est indispensable l’étude du microclimat où vit chaque organisme pour comprendre à quel jeu de facteurs il est effectivement soumis; à son échelle, en effet, les conditions climatiques peuvent être très différentes des caractéristiques générales du climat de la région où il vit.
La loi du minimum , qui est une extension écologique de la loi de Liebig (formulée à propos de cultures expérimentales), découle de la notion de facteur limitant. Elle établit que la présence et le développement complet (jusqu’à la reproduction) d’une espèce dans un milieu donné sont conditionnés par le facteur qui, parmi ceux susceptibles d’être limitants, se trouve le plus proche d’un seuil – minimal ou maximal – de tolérance. Au cours de son développement, l’espèce passe par des stades pouvant présenter des sensibilités différentes aux facteurs du milieu: la présence de l’espèce dans un milieu dépend donc de la valence écologique de son stade le plus sensible , vis-à-vis de ce milieu. On sait que pour les animaux, le stade d’œuf en développement est en général particulièrement sensible, mais c’est souvent le stade larvaire (moins protégé que l’œuf) qui est affecté de la valence écologique la plus basse. Celle-ci augmente généralement au cours du développement, souvent avec des crises répétées (phases de mue, sortie d’hibernation ou d’estivation, puberté, etc.), jusqu’à l’âge sexuellement mûr, écologiquement le plus résistant. Elle diminue à nouveau avec la vieillesse. Des variations cycliques de la valence écologique s’observent en outre chez les espèces pluriannuelles au moment de la reproduction.
Milieu et variations adaptatives des êtres vivants
Chez les végétaux comme chez les animaux, par-delà les divisions taxinomiques nées de l’histoire phylogénétique des groupes, des convergences de morphologie et de physiologie peuvent rapprocher les espèces qui vivent dans des conditions identiques.
Ainsi, on a pu établir chez les animaux supérieurs, sur des bases statistiques, des relations entre certaines caractéristiques physiologiques ou morphologiques et certains facteurs du milieu, en particulier la température. Chez les homéothermes, par exemple, à l’intérieur d’une espèce ou d’un groupe suffisamment homogène d’espèces voisines, les tailles les plus grandes se rencontrent dans les régions les plus froides (règle de Bergmann ). Cela est lié au phénomène de déperdition de chaleur par la surface du corps: proportionnellement plus élevée chez les animaux petits, elle leur rend la régulation thermique plus coûteuse en énergie. Les grands individus sont donc favorisés par la sélection naturelle en milieu plus froid. Cette règle a été confirmée par la paléontologie: lors des glaciations, ce sont les gros animaux qui, parmi les représentants de la faune précédente de climat chaud, ont le plus longtemps persisté, s’adaptant mieux aux climats froids (éléphants, rhinocéros, gros carnivores).
Chez les végétaux, il existe de façon analogue des adaptations à certaines conditions du milieu qui se retrouvent chez diverses espèces dont on dit alors qu’elles appartiennent à la même «forme biologique». Réciproquement, on a été conduit à classer les adaptations différentes que peuvent présenter les diverses espèces soumises aux mêmes contraintes, par exemple le froid. Raunkiaer a proposé ainsi une classification en types biologiques des végétaux de zone tempérée, fondée sur la position des bourgeons par rapport au sol et sur la protection qui leur est ainsi offerte durant la mauvaise saison. Chez les phanérophytes (arbres et arbustes, mais aussi lianes et épiphytes), les bourgeons sont éloignés du sol; chez les chamaephytes (hauts d’au plus 25 cm, par convention), les bourgeons, placés bien plus bas, sont susceptibles d’être protégés par la neige contre les températures hivernales excessives; les hémicryptophytes ont leurs bourgeons juste à la surface du sol, de sorte qu’ils sont protégés même par une faible épaisseur de neige ou simplement par la litière de feuilles mortes; les géophytes, ou cryptophytes, passent la mauvaise saison sous forme de bulbes ou de rhizomes souterrains; les thérophytes, enfin, sont des plantes annuelles qui passent la saison hostile sous forme de graines ou de spores. Cette classification, avec quelques ajustements dans les définitions, peut être transposée à d’autres zones climatiques: il est alors possible de comparer la végétation d’une région à une autre en établissant des spectres biologiques , tableaux précisant la répartition des espèces entre les différents types biologiques.
De telles classifications ont évidemment un caractère statique: un végétal, au cours de son développement, peut se présenter successivement sous différents types, ou au contraire se trouver maintenu dans un certain état, par exemple par le passage d’un feu saisonnier, ou sous l’influence des prédateurs.
Les relations de l’organisme individuel avec le milieu doivent donc être étudiées d’une façon globale et dynamique, chez les végétaux comme chez les animaux, de façon à prendre en compte l’ensemble du cycle vital. Il importe ensuite de comprendre les mécanismes qui assurent le maintien des populations dans le milieu: si les uns jouent à l’échelon des individus en favorisant leur survie, les autres sont mis en jeu à l’échelle des populations.
2. Fonctionnement des populations
Cinétique démographique
Chaque individu d’une espèce naît, vit un certain temps et meurt. Une population est donc un système dynamique dont la permanence résulte de phénomènes compensatoires: des individus quittent le système (mortalité, émigration), tandis que d’autres y pénètrent (natalité, immigration).
L’analyse démographique instantanée d’une population permet de mettre en évidence sa structure, c’est-à-dire sa composition en individus de différentes catégories, en général des classes d’âge.
La détermination de l’âge des individus formant une population naturelle fait appel à des méthodes plus ou moins précises selon les organismes considérés. Lorsque c’est possible, on utilise des caractères morphologiques ou anatomiques traduisant des phases successives de la croissance. Si celle-ci varie périodiquement en relation avec l’alternance des saisons, il peut apparaître des structures comme les anneaux de croissance: de nombreuses plantes ligneuses en présentent et l’on en observe aussi sur les écailles de certains Poissons ou encore sur des coupes transversales d’os chez de nombreux Vertébrés. Souvent, il est seulement possible de distinguer le stade de développement des individus, sans pouvoir leur attribuer un âge précis. C’est le cas pour de nombreux Arthropodes, dont les stades successifs peuvent être reconnus par exemple grâce aux variations numériques de certains organes sensoriels (ocelles, trichobothries) ou de diverses ornementations. Faute de critères d’âge, il reste la possibilité de décomposer la population étudiée en classes de taille, à condition qu’il existe entre la taille et l’âge une relation statistique suffisamment étroite. Dans certains cas, enfin, il est possible de procéder au marquage d’individus d’âge connu et d’en suivre l’effectif au cours d’échantillonnages successifs.
Si l’analyse de la population est poursuivie au cours du temps, elle permet la description des variations temporelles de la structure démographique, c’est-à-dire celles des effectifs des différentes classes reconnues. Ces variations diachroniques constituent ce que l’on appelle la cinétique de la population. Il existe divers modes de représentation des variations temporelles de ces effectifs (tables de survie, matrices de Leslie). La procédure en théorie la plus simple consiste à suivre des ensembles d’individus nés au cours d’un même intervalle de temps, ou cohortes . La courbe représentant l’évolution temporelle de l’effectif d’une cohorte est appelée courbe de survie: sa forme fait ressortir les variations de la mortalité avec l’âge des individus. D’une espèce à l’autre, il existe fréquemment des différences entre les courbes de survie (fig. 4), ce qui traduit des «comportements démographiques» distincts.
L’étude du rythme de succession des cohortes – c’est-à-dire de la répartition temporelle de la reproduction – et l’établissement des courbes de survie correspondantes permettent de décrire le mode de renouvellement d’une population et d’analyser les variations de son effectif total. Il convient alors de connaître le jeu des facteurs, tant biotiques qu’abiotiques, qui détermine la cinétique observée: cette approche causale constitue la dynamique des populations .
Dynamique des populations
Des observations périodiques permettent souvent de constater une relative stabilité des effectifs de nombreuses populations, à condition d’adopter une périodicité d’observation adaptée à la cinétique de chacune d’elles. Il y aurait alors un certain équilibre entre mortalité et natalité, entre émigration et immigration. D’autres fois, en revanche, des pullulations sont observées, ou au contraire des diminutions considérables d’effectifs.
Pour aborder l’étude de ces phénomènes, il est commode de considérer le cas simple d’une population sans émigration ni immigration dont les taux de natalité n et de mortalité m sont constants. On peut alors écrire en appelant N l’effectif de la population et t le temps:

Le paramètre r est appelé taux intrinsèque d’accroissement démographique . S’il est positif, ce qui est habituellement le cas, la population accroît indéfiniment son effectif. Dans la nature, en réalité, toute population atteint plus ou moins rapidement une limite, ce dont rend compte le modèle logistique de Verhulst:

La constance K représente le nombre maximal d’individus que peut comporter la population dans le milieu où elle vit; aussi l’appelle-t-on capacité biotique du milieu pour l’espèce considérée. Bien que trop simple pour représenter la complexité des situations réelles, ce modèle a le mérite de faire ressortir cette idée que l’expansion de toute population est limitée, au sein d’un écosystème, par le jeu de multiples facteurs.
La figure 5 montre les différents types d’interactions qu’une population entretient avec l’écosystème dont elle fait partie: son expansion peut être freinée par les effets limitants de certaines ressources, par les prélèvements opérés par les utilisateurs de la population, par la compétition avec d’autres populations utilisant les mêmes ressources, par les effets, enfin, des facteurs abiotiques jouant à la fois sur les populations elles-mêmes et sur leurs interactions. Des facteurs intrinsèques, c’est-à-dire propres à la population, peuvent également jouer dans la limitation de son effectif.
Ces divers facteurs peuvent provoquer directement la mortalité individuelle et donc influer sur le renouvellement de la population en réduisant le nombre des reproducteurs et leur espérance de vie. De façon plus indirecte, ils peuvent influencer la fertilité et la fécondité, donc le taux de natalité.
La multiplicité des facteurs en jeu rend difficiles aussi bien l’analyse des situations naturelles que l’expérimentation, trop souvent abusivement simplificatrice. L’étude des relations prédateurs-proies en a été un exemple: des modèles différentiels dits «de Lotka-Volterra» ont été développés et vérifiés expérimentalement mais sans doute la vérification ne réussit-elle que dans la mesure où modèles et expériences isolent la relation prédateur-proie de son contexte écologique. En réalité, une population et son environnement forment un système , qui doit être étudié en tant que tel.
Les phénomènes de compétition sont pour cette même raison difficiles à saisir. On dit qu’il y a compétition si plusieurs organismes (de même espèce ou d’espèces différentes) utilisent des ressources communes qui sont en quantité limitée, ou lorsque ces ressources ne sont pas limitantes, si, en les recherchant, les organismes en concurrence se nuisent. La compétition revêt des modalités très variées, notamment selon qu’elle est directe (émission de substances toxiques par un des compétiteurs, par exemple) ou indirecte, lorsque l’un des compétiteurs réduit les ressources exploitables par l’autre.
Pour les organismes fixés, et donc tout spécialement pour les végétaux, la distance entre les individus joue un rôle essentiel. Une proximité trop grande entraîne ainsi une concurrence sévère pour l’eau et les sels minéraux. La compétition pour la lumière est également très importante, et l’on sait les modifications de forme que détermine pour les végétaux la vie en formations denses. Chez les animaux, la compétition présente des aspects comportementaux qui en compliquent l’analyse. On a tenté de reproduire expérimentalement, dans des conditions de milieu bien définies, des phénomènes de compétition entre deux espèces. Les exemples les plus classiques sont ceux des paramécies (Protozoaires Ciliés), des daphnies (Crustacés Cladocères), des Tribolium (Coléoptères des denrées stockées) et des drosophiles (Diptères). Ces différents organismes, tous de petite taille et à reproduction rapide, permettent en effet de suivre un grand nombre de générations dans un volume de milieu réduit. Dans tous les cas, dont la figure 6 représente un exemple, ces expériences de compétition dans un milieu simple conduisent à l’élimination de l’une des espèces en présence. Dans certaines circonstances, on peut toutefois inverser les résultats en modifiant les conditions du milieu, par exemple l’alimentation. C’est seulement en rendant le milieu plus complexe qu’il est possible d’obtenir la coexistence durable des deux espèces, qui exploitent alors des parties différentes de leur environnement.
La compétition peut être interspécifique, mais aussi intraspécifique: on conçoit que celle-ci devient d’autant plus probable que l’effectif de la population augmente. Comme le suggère le modèle logistique, l’accroissement d’une population dans un espace fini doit provoquer la mise en jeu de mécanismes limitant cette expansion, mécanismes dits densité-dépendants .
La compétition intraspécifique, pour être bien comprise, doit être replacée dans une perspective évolutionniste: ses modalités actuelles sont le fruit de la sélection naturelle et il faut tenter d’en comprendre la signification adaptative. Dans cette optique, il apparaît de plus en plus que les interactions sociales jouent un rôle dans la régulation des effectifs, par exemple lors des phénomènes de territorialisation: on sait que chez bien des espèces, la taille des territoires est d’autant plus importante que les ressources sont moins abondantes.
En fait, la compétition intraspécifique est dans une large mesure à l’origine de la sélection naturelle: ce sont les compétiteurs les plus efficaces qui participent le plus au renouvellement de la population et donc à la composition du pool génique: ici s’imbriquent la dynamique des populations, l’éthologie et la génétique en une problématique qui constitue l’essentiel de la sociobiologie .
Le rôle des phénomènes densité-dépendants dans la régulation des populations naturelles a fait un temps l’objet de très vives controverses, de nombreux auteurs admettant que la limitation des effectifs est due davantage aux facteurs physiques et à leurs variations parfois brutales. La prise en compte du concept de «système population-environnement » permet de dépasser cette opposition. Il s’est développé actuellement une théorie générale de la régulation des populations intégrant les facteurs extrinsèques (biotiques et abiotiques) et intrinsèques (comportements en particulier). Cette théorie, au lieu de plaider la prépondérance générale d’un type de mécanisme sur les autres, s’attache à distinguer différents types de populations en fonction de la part relative des mécanismes indépendants de la densité et de celle des facteurs qui en dépendent. Elle cherche ensuite à interpréter les différences de fonctionnement démographique dans une perspective évolutionniste.
Concept de niche écologique
Dans un écosystème, la population d’une espèce donnée entretient de nombreuses interactions avec les autres composantes de la biocénose en même temps qu’elle est soumise aux caractéristiques particulières de l’espace qu’elle occupe dans le biotope. Comparée aux populations d’autres espèces vivant dans le même écosystème, cette population a donc avec celui-ci un ensemble de relations qui lui est propre et qui constitue sa niche écologique .
Ce concept a fait l’objet de discussions nombreuses et il n’a pas toujours eu le caractère «opératoire» d’un véritable concept scientifique. Bien que le terme français de niche fasse surtout penser à la simple localisation spatiale d’un organisme individuel dans son milieu, il paraît préférable d’accorder au concept une signification plus complète: la niche d’une espèce désigne non seulement le type de place qu’occupent dans le milieu les individus de cette espèce, mais aussi les multiples relations qu’ils ont avec le reste de l’écosystème; en d’autres termes, la niche écologique d’une espèce serait dans un écosystème à la fois son habitat et sa fonction.
L’expérience et l’observation s’accordent pour montrer que deux espèces n’occupent jamais exactement la même niche, même dans un écosystème complexe. Réciproquement, on peut dire que deux espèces qui coexistent ont certainement deux niches différentes, cette différence pouvant n’être que partielle et ne porter que sur une caractéristique déterminée comme le cycle saisonnier, le rythme nycthéméral, les proies habituelles, etc.
Il est illusoire d’espérer caractériser dans sa totalité la niche écologique d’une population: ce serait savoir tout sur elle et sur l’écosystème dont elle fait partie. En revanche, il est essentiel de pouvoir comparer entre populations différentes certaines composantes de leurs niches respectives et d’apprécier ainsi les conditions réelles de leur coexistence. On pourra par exemple étudier la dimension trophique de ces niches, c’est-à-dire comparer des régimes alimentaires afin de déterminer quelles ressources sont susceptibles de faire l’objet d’une compétition interspécifique.
Il y a ainsi un lien étroit entre le concept de niche et la notion de ressources qui évoque la disponibilité de l’écosystème vis-à-vis de tel ou tel besoin précis de la population. Celle-ci, en effet, utilise un certain espace à l’intérieur duquel elle doit trouver l’énergie et les constituants chimiques nécessaires à l’entretien et au renouvellement des individus qui la composent. Ces besoins sont à la fois quantitatifs et qualitatifs, et toute ressource correspondante est susceptible de se révéler limitante; il est donc important de les mesurer en établissant des bilans à l’échelle des populations, comme par exemple le bilan énergétique.
Bilan énergétique des populations
La connaissance de la consommation d’aliments, celle de la production et le calcul des bilans énergétiques amènent à mieux comprendre le rôle des populations au sein des écosystèmes où elles vivent. Comme un individu, une population est caractérisée par les quantités d’énergie qu’elle ingère (I), qu’elle assimile (A), ou excrète (NA), qu’elle respire (R) et qu’elle transforme en nouvelle matière vivante (P). Ces quantités sont rapportées à l’unité de temps – généralement l’année – et établies pour la population entière ou un échantillon représentatif de celle-ci. De même qu’à l’échelle individuelle, on définit les rapports P/I (rendement écologique de croissance), P/A (rendement de croissance tissulaire) et A/I (rendement d’assimilation).
Au cours d’un cycle annuel, la population a consommé, dépensé et produit les sommes des consommations, dépenses et productions des divers individus qui en ont fait partie. Établir le bilan énergétique de la population, c’est par conséquent combiner la connaissance des bilans individuels, le plus souvent mesurés en élevage, et celle des variations de la structure de la population, suivie sur le terrain. Cette intégration est facilitée par l’existence fréquente au sein d’une espèce de relations quantitatives entre les différentes composantes du bilan individuel et la taille ou la biomasse de l’organisme.
La production d’une population peut être calculée à l’aide de diverses techniques. L’une des plus simples est l’utilisation de la courbe d’Allen qui permet de déduire graphiquement la production d’une cohorte d’individus de la relation entre l’effectif et le poids individuel à un moment donné (fig. 7). Évidemment, cette technique suppose que la population puisse être décomposée en cohortes bien définies.
Chez l’individu, le rendement d’assimilation est très lié au régime alimentaire et varie donc assez peu au cours de sa vie, sauf s’il y a des changements importants de régime au cours du temps. À l’échelle de la population, le rapport A/I conserve donc habituellement une valeur très proche de celle correspondant à l’individu.
La production individuelle variant avec l’âge, les valeurs des rendements P/A et P/I, à l’échelle de la population sont liées à la cinétique démographique. D’une façon générale, le rendement P/A varie de 0,30 à 0,10 chez les Invertébrés hétérothermes herbivores et détritivores, ainsi que chez les Vertébrés hétérothermes carnivores. Il est bien plus faible (0,05 à 0,01) chez les homéothermes.
Le rendement écologique de croissance P/I est également très variable: toujours faible chez les Vertébrés homéothermes, ainsi que chez les Invertébrés détritivores, il est en revanche relativement élevé chez les hétérothermes carnivores (Vertébrés et Invertébrés), moyen chez les Invertébrés hétérothermes herbivores (tableau).
Les rapports P/I, P/A et A/I traduisent des rendements énergétiques. Pour comprendre le rôle de la production des diverses populations dans le fonctionnement d’un écosystème, il convient de rapporter cette production P à la biomasse B qui lui donne naissance. La production étant mesurée pendant un intervalle de temps déterminé, on considère son rapport à la biomasse moyenne 祿 de la population pour l’intervalle considéré, soit P/ 祿, qui représente le taux de renouvellement de la biomasse et qui, au contraire des rendements énergétiques, fait donc intervenir le facteur temps.
La valeur de P/ 祿, apparaît liée essentiellement à la durée de vie des organismes: de l’ordre de 3 à 5 chez les animaux dont la longévité maximale est de l’ordre d’une année, elle est plus élevée chez ceux dont l’existence ne dure que quelques mois et ont plusieurs générations par an; elle est plus faible au contraire chez ceux qui vivent plusieurs années, descendant ainsi à 0,3 pour le daim qui peut atteindre une dizaine d’années et à 0,05 pour l’éléphant (qui dépasse souvent 60 ans).
L’inverse du taux de renouvellement 祿/P représente le temps de renouvellement de la biomasse, c’est-à-dire en principe la durée de vie moyenne d’un individu.
Chez les végétaux ligneux, la signification de P/ 祿 est particulière, car une part importante de la production est souvent perdue chaque année par l’individu, sous forme de feuilles, de rameaux et de radicelles. À l’échelle de la vie de l’individu, l’accroissement de biomasse B est ainsi inférieur à la production. Dans ces conditions, le rapport P/ 祿 ne représente pas le taux de renouvellement de la biomasse sur pied et son inverse est très inférieur à la durée de vie moyenne des individus. Pour une forêt de chênes ainsi que pour un bois de pins pignons, il a été trouvé une valeur de 0,09 environ pour P/ 祿, soit onze ans pour 祿/P, alors que la durée de vie moyenne de ces arbres est bien plus élevée.
Pour les espèces herbacées pérennes, le rapport P/ 祿 est de l’ordre de 1 pour l’année. Il est supérieur à 1 pour les espèces herbacées annuelles. Les valeurs de P/ 祿 sont naturellement bien plus fortes encore pour les espèces formant le phytoplancton: calculées pour les périodes les plus favorables du cycle annuel, elles peuvent largement dépasser 100.
Le rapport P/ 祿, on le voit, a l’intérêt de caractériser de façon très synthétique le rôle de chaque population dans l’élaboration de matière vivante au sein de l’écosystème. En outre, lorsqu’il est par avance connu avec une précision suffisante – ce que permet souvent la détermination de la cinétique démographique –, il permet d’estimer la production d’une population à partir de la biomasse moyenne de celle-ci.
3. Organisation fonctionnelle des biocénoses
Organisation trophique
Un des problèmes majeurs de l’écologie actuelle est la description et la compréhension de la structure trophique des principaux écosystèmes.
Cette structure trophique est constituée par les relations qui lient les organismes consommés à ceux qui les consomment, relations dont l’ensemble forme le réseau trophique du système. Ce terme traduit bien mieux que celui de chaînes alimentaires les relations innombrables qui existent entre les différentes espèces de la biocénose.
Selon la source d’où ils tirent leur énergie, les êtres vivants se répartissent entre deux grandes catégories. Les autotrophes édifient leur matière organique à partir de constituants minéraux où dominent l’eau, le gaz carbonique, des nitrates et des phosphates; le mécanisme de loin le plus répandu de cette élaboration est la photosynthèse, réalisée grâce à l’apport énergétique du rayonnement lumineux capté à l’aide de la chlorophylle présente chez les végétaux supérieurs. Diverses Bactéries également autotrophes utilisent d’autres sources d’énergie, diverses substances minérales notamment. Les hétérotrophes utilisent la matière organique ainsi formée et se bornent à la transformer. Ils sont représentés par l’ensemble des animaux, mais aussi par beaucoup de Bactéries et par des végétaux non chlorophylliens comme les Champignons.
L’établissement du réseau trophique d’un écosystème est une tâche difficile par suite du grand nombre d’espèces en présence et de la complexité du régime alimentaire de chacune. Non seulement, en effet, un individu se nourrit généralement, à un moment donné de son existence, aux dépens de nombreuses autres espèces, mais son spectre de nourritures varie avec son stade de développement, son âge, sa taille, et aussi avec la période de l’année et les disponibilités qu’elle offre.
Si simple que soit une biocénose, le réseau trophique complet est ainsi d’une complexité telle qu’il importe, pour en comprendre l’essentiel, de le présenter sous une forme plus simple et de le schématiser. Pour cela, on conserve seulement les espèces les plus abondantes et surtout on regroupe les espèces d’un même groupe taxinomique dont les exigences trophiques sont voisines: par exemple les vers de terre géophages, ou l’ensemble des araignées (fig. 8). Même ainsi, cependant, les relations sont encore complexes et l’on pourra ne considérer qu’une fraction de la biocénose, par exemple les organismes du sol, ou encore ceux de la strate herbacée. On considère parfois aussi dans le réseau trophique la partie qui est en relation avec une espèce particulière à laquelle on s’intéresse plus spécialement.
Donner d’un réseau trophique une idée quantitative en précisant les biomasses des divers ensembles indiqués constitue un idéal difficile à atteindre. De fait, peu de biocénoses ont permis d’arriver à un tel résultat. Aussi a-t-on tenté de simplifier davantage en n’y considérant que quelques ensembles synthétiques auxquels on a donné le nom de niveaux trophiques.
Le premier niveau est celui des producteurs encore appelés producteurs primaires , qui regroupe essentiellement les végétaux chlorophylliens. À l’origine de toutes les chaînes trophiques, ils sont consommés par des herbivores (ou phytophages) qui constituent le second niveau, celui des consommateurs primaires. Ceux-ci servent à leur tour d’aliments à des carnivores (ou parasites) formant le niveau des consommateurs secondaires. Beaucoup d’auteurs considèrent également un niveau supplémentaire de consommateurs tertiaires , carnivores consommant des carnivores.
En réalité l’individualité de ces niveaux de consommateurs est toute relative, car un très grand nombre d’animaux sont plus ou moins omnivores, ou tout au moins polyphages: leur régime comprend à la fois des végétaux et d’autres animaux, herbivores ou carnivores, de sorte que beaucoup d’espèces se situent simultanément à plusieurs niveaux.
Une autre difficulté de l’établissement des niveaux trophiques est liée à l’existence des micro-organismes, Bactéries et Champignons, dont la place dans le réseau trophique est en général plus difficile à préciser. Aussi les a-t-on souvent classés dans un niveau trophique particulier, le niveau des décomposeurs. Il n’existe pourtant pas de différence de nature entre le rôle trophique des micro-organismes décomposeurs de matière organique et celui des saprophytes et nécrophages macroscopiques, non plus qu’entre le régime de ceux-ci et celui des vrais herbivores ou prédateurs. Seul leur pouvoir élevé de consommation, lié à une activité métabolique intense, donne une impression d’évanouissement rapide de la matière qu’ils consomment, à l’opposé des organismes de plus grande taille, à croissance plus lente. Ils sont aussi bien moins connus et étudiés.
Ajoutons que des interactions complexes existent entre les micro-organismes et beaucoup d’animaux dits détritivores qui, en fait, se nourrissent des micro-organismes. Certains les consomment dans le milieu extérieur, d’autres les élèvent (termites et fourmis champignonnistes), d’autres encore digèrent progressivement ceux qui se développent dans leur tube digestif (termites).
D’autres interactions sont liées seulement au rôle mécanique de nombreux détritivores qui, en triturant les débris animaux et surtout végétaux, en facilitent l’attaque par les Bactéries. Ils bénéficient à leur tour de cette digestion partielle qui leur procure des aliments plus accessibles à leurs enzymes digestives.
Que les micro-organismes décomposeurs soient intégrés ou non dans les niveaux trophiques classiques, ceux-ci sont souvent représentés graphiquement par une pyramide, dite pyramide des biomasses , dont les étages successifs correspondent aux niveaux qui viennent d’être définis (fig. 9). La surface de chaque étage de la pyramide est proportionnelle à la biomasse du niveau qu’il représente. On a naturellement intérêt, pour faciliter les comparaisons, à exprimer cette biomasse par son équivalent énergétique en calories ou en joules, qui, compte tenu de la présence des squelettes externes ou internes, des coquilles, des inclusions minérales diverses, traduit mieux que le poids sec l’importance réelle de la matière vivante des différents groupes d’êtres vivants.
Dans les biocénoses étudiées en détail, il est possible de faire ressortir les parts respectives des productions endogées et épigées, ainsi que celles des micro-organismes par rapport à celles des macro-organismes. Signalons aussi que l’usage d’une échelle logarithmique facilite souvent la présentation.
Le réseau trophique comme la pyramide des biomasses ne donnent de l’organisation trophique qu’une image purement statique. Ce sont les transferts d’énergie d’une espèce à l’autre, ou d’un niveau trophique à l’autre (et par suite les flux d’énergie dans l’écosystème), qu’il faut quantifier pour en décrire le fonctionnement, reprenant ainsi pour l’ensemble du système les bilans d’énergie décrits pour les populations. De tels transferts sont définis par leur débit , qui est la quantité d’énergie transférée dans un temps donné, et aussi par leur rendement , rapport de l’énergie réellement utilisée à celle qui a été prélevée.
Peuplements et guildes
La complexité des biocénoses est souvent telle qu’on ne peut d’emblée en faire l’étude complète: il faut les décomposer en sous-ensembles constituant des niveaux d’organisation intermédiaire entre la population et la biocénose.
Ces sous-ensembles sont généralement délimités pour des raisons méthodologiques d’observation et d’échantillonnage; fréquemment, cela revient à étudier des populations d’espèces d’un même groupe taxinomique, plus ou moins large: selon les circonstances, on étudiera par exemple tous les Insectes du milieu herbacé, ou seulement les Orthoptères, ou même les seuls criquets.
On peut désigner par le terme de peuplement ces divers sous-ensembles délimités plus ou moins arbitrairement au sein d’une biocénose. Un peuplement défini sur des bases taxinomiques a l’avantage de pouvoir être étudié par des chercheurs spécialisés dans la systématique du groupe retenu. Une telle spécialisation de nature taxinomique, qui a marqué une grande partie des recherches écologiques, n’est toutefois pas sans danger car elle risque de conduire à un cloisonnement, masquant le but majeur de l’écologie qui est de comprendre le fonctionnement d’ensemble de la communauté des êtres vivants d’un milieu. Ce risque est d’autant plus grand que le terme de peuplement est particulièrement vague.
Aussi est-il souvent plus efficace de prendre en considération des sous-ensembles qui soient définis non seulement pour des raisons pratiques, mais aussi parce qu’ils ont une signification fonctionnelle précise. Pour cette raison les recherches portent de plus en plus sur des groupes de populations appartenant à des espèces taxinomiquement apparentées et qui exploitent, au sein de l’écosystème, le même type de ressources. On étudiera ainsi les populations d’Oiseaux frugivores, les larves de Diptères saprophages, les araignées installant des toiles de capture dans la strate herbacée, les petits Mammifères granivores... Les sous-ensembles définis de cette manière sont parfois appelés des guildes (fig. 10).
La notion de guilde, plus précise que celle de peuplement, n’en comporte pas moins une part d’arbitraire. En premier lieu, le type de ressources par rapport auquel elle est définie peut être délimité de façon plus ou moins étroite. En second lieu, une même ressource peut être exploitée par des populations sans lien systématique: dans une forêt tropicale, par exemple, les fruits sont consommés non seulement par des Oiseaux, mais aussi par des chauves-souris et des Primates, ainsi que par des Insectes. À la limite, surtout lorsque la ressource considérée n’est pas répartie de façon diffuse dans l’écosystème mais forme des entités spatialement bien délimitées, comme une espèce d’arbuste dans une savane ou des troncs morts dans une forêt, il est important d’étudier l’ensemble des populations qui l’exploitent à un titre ou à un autre: on peut parler alors de la synusie d’un arbuste, des arbres morts, etc.
Dans tous les cas, l’étude de peuplements, de guildes ou de synusies vise dans un premier temps à mettre en évidence leur structure, puis à en rechercher une interprétation fonctionnelle. Cette structure peut être étudiée sous quatre aspects: la composition spécifique, la répartition spatiale, la distribution temporelle des cycles biologiques et des activités, la structure en taille.
La façon la plus simple de caractériser la structure spécifique est évidemment le nombre d’espèces en présence qu’on appelle aussi richesse spécifique. Cette notion de richesse présente toutefois deux inconvénients. Le premier, purement pratique, c’est qu’il est difficile, dans un milieu, de connaître le nombre total d’espèces, même d’un groupe limité: certaines sont rares et risquent d’échapper à l’échantillonnage. Dans les faits, la richesse est une fonction croissante de la qualité de l’échantillonnage. L’autre inconvénient, d’ordre théorique, c’est que la richesse ne tient pas compte de l’abondance relative des espèces. Aussi a-t-on introduit la notion de diversité spécifique qui tient compte de ces différences de fréquence. L’indice le plus utilisé pour exprimer cette diversité, appelé indice de Shannon, est lié à la théorie de l’information. Il s’écrit:
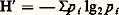
où p i est la proportion de l’espèce de rang i , les diverses espèces ayant été classées par ordre de fréquence décroissante. Pour un nombre d’espèces donné, H est maximal lorsque toutes les espèces ont la même fréquence.
La valeur de l’indice de Shannon dépend entre autres de la superficie considérée. Au sein d’une biocénose bien définie, on l’appelle H size=1見. Dans un complexe écologique plus vaste et plus diversifié – ensemble de biocénoses –, c’est H size=1塚.
De façon plus complète que sa richesse et sa diversité, c’est la loi de distribution des abondances des diverses espèces qui définit la structure spécifique d’un peuplement. Elle est représentée graphiquement en portant en ordonnée le nombre d’individus vivant sur une superficie déterminée et en abscisse le rang des espèces classées par abondance décroissante (fig. 11). On voit immédiatement apparaître l’évidente nécessité de ne considérer ainsi qu’un ensemble de formes présentant un minimum d’homogénéité, ne serait-ce que de taille.
Selon les peuplements, les distributions d’abondance des espèces peuvent être comparées à divers modèles théoriques dont certains traduisent des hypothèses écologiques relatives au partage des ressources.
Le modèle de Motomura correspond à une série géométrique, c’est-à-dire à de très fortes inégalités entre les fréquences des espèces: K, K(1 漣 K), K(1 漣 K)2, ... K(1 漣 K)n . Il représente bien des communautés simples, où une très forte compétition ne permet la coexistence que d’un nombre limité d’espèces dont un très petit nombre seulement est largement dominant.
Le modèle de Preston, ou distribution dite log-normale, s’applique surtout, au contraire, à des situations où agissent de nombreux facteurs et où coexistent de nombreuses espèces. C’est une distribution de fréquence qui prend la forme d’une distribution normale quand on transforme la variable en son logarithme. À l’encontre du modèle de Motomura, la majorité des espèces présente des abondances moyennes.
De tels modèles de distribution d’abondance doivent être interprétés avec précaution. Leur intérêt principal est de permettre de comparer des stades successifs d’évolution d’un peuplement ou encore divers peuplements d’un même groupe taxinomique dans diverses biocénoses. D’un groupe à un autre, cependant, la comparaison risque d’être faussée par des différences entre les richesses des groupes. C’est pourquoi ces méthodes descriptives ne dispensent pas d’une analyse fonctionnelle.
Relations interspécifiques
Les flux de composés chimiques et d’énergie circulant d’une espèce à l’autre constituent l’aspect majeur du fonctionnement des biocénoses. La compréhension de celui-ci suppose cependant qu’au-delà de la simple description des voies de circulation de la matière et de l’énergie, on saisisse le jeu des relations interspécifiques qui la règlent. En effet, tout flux trophique établi entre deux populations est quantitativement déterminé par la dynamique de chacune d’elles, donc notamment par les relations entre les populations en présence.
Au sein d’une biocénose, il y a trois types fondamentaux d’interactions entre espèces. L’exploitation d’une population par une autre correspond aux rapports entre prédateurs et proies, animales ou végétales, aussi bien qu’entre parasites et hôtes, toutes les transitions existant entre ces deux types. La compétition résulte du fait qu’en exploitant une même catégorie de ressources deux ou plusieurs populations se nuisent mutuellement (cf. supra ). Au contraire, lorsqu’il y a symbiose , les partenaires tirent chacun bénéfice de l’activité des autres.
Exploitation
Les rapports entre les végétaux et les animaux phytophages mettent souvent en jeu des substances chimiques, élaborées par les plantes, qui ont un rôle de défense ou sont au contraire attractives. Les substances de défense peuvent avoir une simple action répulsive, mais d’autres sont de véritables insecticides. Certaines plantes élaborent des molécules qui sont mimétiques des hormones des Insectes, et peuvent ainsi en perturber le développement. Les substances attractives déterminent généralement une certaine spécificité de la relation plante-insecte: elle est très étroite dans le cas d’espèces monophages, ou peut concerner plus largement une famille de plantes et une famille d’Insectes.
Le déterminisme des rapports de prédation chez les animaux est très variable. Il existe des prédateurs étroitement spécialisés à propos desquels se pose le problème des mécanismes de repérage des proies, de même que se pose celui de la découverte des hôtes par leurs parasites spécifiques. Il est possible, ici aussi, que la spécificité des relations soit assurée par des interactions chimiques.
Les proies possèdent souvent des moyens de défense chimiques, tels certains termites élaborant des substances de défense contre les fourmis, ou de nombreux mille-pattes (Diplopodes) qui produisent par exemple de l’acide cyanhydrique. Dans ces cas, la défense a lieu à l’échelle individuelle. Dans d’autres, les animaux contiennent des substances toxiques ou répulsives qui ne sont pas émises à l’extérieur: elles n’ont d’effet que si l’individu est ingéré et leur efficacité n’est donc réelle que si le prédateur est capable d’apprentissage. De fait, les animaux contenant de telles substances – qui proviennent souvent des végétaux consommés – possèdent fréquemment des traits d’ornementation remarquables: couleurs vives et contrastées, dessins très simples. On pense que ces caractères peuvent être mémorisés par certains prédateurs (Oiseaux, Mammifères).
Bien des prédateurs sont plutôt «généralistes», consommant des proies variées. Cependant, il existe souvent une relation assez précise entre la taille d’un prédateur et celle de ses proies. Cette relation dépend évidemment des caractéristiques morphologiques du prédateur, mais peut-être aussi d’un rapport optimal entre l’apport énergétique que représente une proie d’une certaine taille et l’énergie dépensée pour sa capture.
Compétition
Lorsque des guildes peuvent être délimitées dans une biocénose, il se pose le problème de savoir si les populations qui les constituent sont en compétition effective pour l’exploitation de leurs ressources communes et dans quelle mesure cette compétition détermine la répartition des flux trophiques.
On constate souvent que les ressources font l’objet d’un partage, par exemple dans l’espace ou encore dans le temps: cela réduit les risques de compétition interindividuelle, mais n’explique pas toujours la limitation numérique des populations potentiellement compétitrices. En fait, la compétition n’intervient que si les besoins trophiques des populations exploitantes dépassent la capacité de production des populations exploitées. Lorsque les premières sont elles-mêmes l’objet d’une limitation par des prédateurs ou des parasites, cette situation peut ne pas se produire.
Lorsque la catégorie de ressources considérée est relativement hétérogène, il arrive que les espèces de la guilde soient spécialisées dans l’exploitation d’une partie seulement des ressources. Cette spécialisation se fait souvent par rapport à la taille: ainsi une guilde d’Oiseaux granivores peut-elle comprendre plusieurs espèces se nourrissant de graines de tailles différentes, en fonction de leur grosseur. De tels constats soulèvent le problème des processus évolutifs ayant conduit aux situations observées: la spécialisation est-elle le fruit d’une sélection induite par le jeu effectif de la compétition interspécifique? Celle-ci aurait alors joué le rôle d’un facteur sélectif diversifiant, tandis qu’elle ne fonctionnerait plus que comme facteur normalisant une fois la spécialisation établie.
Symbiose
La symbiose, ou mutualisme , est un phénomène très répandu dans la nature et qui y joue un rôle souvent essentiel.
Dans le monde végétal, les Lichens sont, on le sait, formés par l’association permanente de Champignons et d’Algues vertes microscopiques. Chez beaucoup de Végétaux supérieurs, la nutrition est favorisée par la présence de Champignons associés aux racines (mycorhizes); des Bactéries nitrifiantes vivent dans les tissus radiculaires des Légumineuses et d’autres espèces végétales.
Beaucoup d’animaux abritent dans leurs tissus des Algues unicellulaires microscopiques qui, par leur photosynthèse, apportent à leurs hôtes un appoint non négligeable d’aliments. Ces Algues sont les Zoochlorelles et les Zooxanthelles, présentes notamment chez certains Protozoaires, des Spongiaires, des Cœlentérés.
Certaines Bactéries et des Levures habitent en permanence l’intestin d’Insectes, souvent en des emplacements spéciaux. Des Levures sont surtout abondantes chez les xylophages. De même, des Flagellés permettent à certains termites la digestion de la cellulose. Des Bactéries et des Protozoaires sont présents dans l’intestin des Vertébrés, notamment chez ceux qui ont un régime essentiellement cellulosique; on sait leur importance dans la digestion des Ruminants et des Équidés.
Ces cas de mutualisme sont relatifs à des relations de nature trophique. Il en est d’autres qui concernent en même temps les mécanismes de la reproduction: certaines plantes, en effet, ne peuvent être pollinisées que par des Insectes qui collectent leur pollen, parfois par des Mollusques ou même des chauves-souris. Dans certains cas, il a été démontré au sein de guildes d’Insectes butineurs (bourdons) une spécialisation morphologique: la longueur de la trompe y varie d’une espèce à l’autre en relation avec la profondeur des fleurs préférentiellement fréquentées. La relation plante-insecte peut être plus subtile encore: c’est ainsi que les Orchidées du genre Ophrys attirent certains mâles d’Hyménoptères par l’émission d’un mélange de substances imitant la phéromone sexuelle des femelles: ces mâles, en tentant de s’accoupler avec les fleurs, en réalisent la pollinisation qui ne peut pas avoir lieu autrement.
Qu’il s’agisse de relations d’exploitation, de compétition ou de symbiose, il existe entre les espèces des coadaptations étroites, mettant souvent en jeu des substances chimiques spécifiques. On a donné à celles-ci le nom d’écomones , en désignant ainsi toute substance (ou complexe de substances) produite par les individus d’une espèce et induisant chez ceux d’autres espèces des effets spécifiques. L’extraordinaire variété de ces coadaptations, leur non moins extraordinaire précision, posent le problème des processus évolutifs qui ont conduit à leur mise en place.
4. Stabilité, adaptabilité et évolution des biocénoses
Complexité et stabilité
À l’échelle de l’observation humaine, de nombreuses biocénoses paraissent stables: leurs caractéristiques structurales et dynamiques ne changent pas sensiblement d’une année à l’autre. Cette permanence résulte, comme pour tout système biologique, d’un équilibre dynamique entretenu par les flux de matière et d’énergie qui traversent le système.
Bien souvent, cependant, l’environnement d’une biocénose présente des irrégularités temporelles qui ne sont pas des variations cycliques normales: ce sont des perturbations que la biocénose répercute dans son fonctionnement. À chaque fois, elle est ainsi écartée de son état normal, mais est susceptible d’y revenir lorsque cesse la perturbation. Les mécanismes de régulation assurant ce retour peuvent être plus ou moins efficaces et l’on est donc conduit à étudier la capacité de stabilisation d’une biocénose en fonction de l’amplitude des perturbations qu’elle peut subir.
De manière générale, les variations de l’environnement ont pour conséquence la modification des cinétiques démographiques de certaines espèces, dont les effectifs sont ainsi augmentés ou diminués de façon inhabituelle. Les transferts trophiques auxquels participent ces espèces sont donc perturbés, ce qui peut avoir des conséquences en cascade au long des chaînes alimentaires. En fait, il n’en est pas toujours ainsi, car l’organisation en réseau de la biocénose permet des compensations interspécifiques: si par exemple l’une des proies habituelles d’un prédateur devient déficitaire à la suite d’une perturbation du milieu, le prédateur peut reporter ses efforts de chasse sur une autre espèce.
Le comportement homéostatique de la biocénose pourrait ainsi être lié à une certaine souplesse du fonctionnement, due au fait que des espèces différentes peuvent remplir des fonctions équivalentes au sein du réseau trophique. La capacité de stabilisation d’une biocénose dépendrait alors de la complexité de celle-ci: plus la biocénose serait riche en espèces, plus le réseau trophique serait complexe et moins le fonctionnement risquerait d’être enrayé du fait de perturbations fortuites.
Cette conception très répandue est toutefois discutable, car ni les recherches théoriques ni les observations dans la nature ne lui donnent de fondements certains; on sait par exemple qu’il est des écosystèmes qui, bien que plus simples que d’autres, se révèlent plus résistants aux perturbations. La discussion résulte en fait de la relative imprécision des concepts de stabilité et de complexité. Le vrai problème est de comprendre quelles propriétés structurales et fonctionnelles permettent à une biocénose de survivre aux effets de perturbations plus ou moins importantes.
Dans le cas d’une biocénose subissant autour d’un état stationnaire moyen des fluctuations ne dépassant pas sa capacité de stabilisation (fig. 12), un accident important peut provoquer la disparition ou le glissement vers un autre état stationnaire. Celui-ci est caractérisé par une organisation différente où la structure spécifique et le réseau trophique sont modifiés qualitativement et quantitativement. Cela peut se produire à la suite de perturbations passagères, suffisantes néanmoins pour provoquer l’acquisition rapide d’un nouvel état capable de se maintenir même après retour du milieu aux conditions initiales, mais peut se produire aussi par adaptation, la biocénose s’ajustant cette fois à un contexte définitivement transformé. Il faut donc très soigneusement distinguer d’une part la capacité de stabilisation, d’autre part la capacité d’adaptation : à la suite de modifications momentanées, une biocénose peut manifester des moyens de retour à l’état initial très puissants, sans être pour autant capable de se transformer avec une efficacité suffisante pour s’adapter à un milieu durablement modifié. On définira donc l’adaptabilité d’une biocénose comme sa capacité à s’ajuster aux changements définitifs de l’environnement par modification de sa structure et de son fonctionnement.
Notion de stratégie adaptative
Une biocénose est un assemblage de populations qui sont elles-mêmes des assemblages d’individus: on ne peut comprendre comment l’organisation d’une biocénose s’adapte aux modifications du milieu sans saisir ce qui se passe aux niveaux d’intégration inférieurs.
L’ensemble des moyens permettant à un système biologique – individu, population, biocénose – d’assurer sa survie dans un environnement susceptible de se transformer, constitue ce qu’il est convenu d’appeler sa stratégie adaptative. Il s’agit de moyens actuels , fruits d’une évolution passée , déterminant pour le futur l’aptitude du système à survivre.
Un individu possède des moyens de s’adapter déterminés par son patrimoine génétique; ils sont à la fois comportementaux et physiologiques et constituent donc ce que l’on appelle une stratégie éthophysiologique. L’individu ne peut évidemment changer son génome: il s’adapte dans la mesure de ses capacités physiologiques de régulation et dans la mesure des moyens comportementaux lui permettant de se soustraire à des conditions défavorables.
La survie d’une population dépend à la fois des stratégies des individus qui la composent et de la façon dont ils sont renouvelés: l’ensemble de ces moyens constitue la stratégie démographique de la population. Vis-à-vis d’une population, un changement de l’environnement se traduit par un nouveau contexte sélectif: il y a sélection d’individus en partie différents de ceux que retenait le contexte sélectif antérieur, à condition évidemment que ces nouveaux individus soient produits par la population. La capacité de celle-ci à réaliser un brassage génique important et rapide joue donc un rôle essentiel: elle est d’autant plus grande que l’espèce se reproduit plus souvent et en plus grand nombre. Il s’agit donc d’une caractéristique de la population, système autoproducteur, qui, avec les moyens propres aux individus, concourt à la survie de l’espèce.
Il existe ainsi des populations dont les individus ont des stratégies éthophysiologiques peu efficaces: leur taux de mortalité est donc élevé, mais il est compensé par une reproduction rapide et abondante. D’autres espèces, au contraire, se renouvellent lentement, mais les individus ont des stratégies éthophysiologiques élaborées qui leur confèrent une probabilité de survie élevée. Tous les intermédiaires existent entre ces deux situations qui font ressortir l’importance de la combinaison des moyens individuels et démographiques vis-à-vis de la survie des populations.
Comme tout autre système biologique, une biocénose est globalement confrontée à des problèmes de survie; ceux-ci mettent en jeu, en plus des stratégies éthophysiologiques et des stratégies démographiques des individus et des populations, des mécanismes propres au système intégré que forme la biocénose.
L’ensemble de ces moyens superposés correspondant aux différents niveaux d’intégration constitue la stratégie cénotique de la biocénose, d’autant plus élaborée que celle-ci possède davantage de mécanismes assurant la régularité des transferts trophiques.
Lorsque l’environnement se transforme, la survie d’une biocénose simple, où certaines fonctions ne sont assurées que par une espèce, n’est possible que si toutes les espèces correspondantes sont capables de s’adapter aux nouvelles conditions de milieu. Sinon les transferts trophiques qu’elles effectuent s’arrêtent, faute d’espèces capables de les relayer. L’existence de formes de rechange favorise au contraire le maintien des flux de matière et d’énergie.
Certes, la coexistence de populations remplissant exactement la même fonction en même temps n’est sans doute pas possible. Cependant, si l’on conçoit la niche d’une espèce comme l’ensemble des fonctions que celle-ci est susceptible de remplir, la coexistence de formes écologiquement proches est possible si chacune ne remplit effectivement qu’une fonction qui lui est propre. Si l’une des espèces vient à disparaître, une autre pourra la remplacer dans l’accomplissement de sa fonction, par exemple en élargissant son régime alimentaire. Il apparaît ainsi que la stratégie cénotique d’une biocénose dépend tout particulièrement de la richesse spécifique des différentes guildes qui la composent et de l’importance du «recouvrement» des niches potentielles des espèces de chaque guilde. Le problème se pose alors de savoir comment se fait, au cours de l’évolution, l’enrichissement des biocénoses en espèces.
Formation des biocénoses et coévolution
L’augmentation du nombre des espèces d’une biocénose est liée en premier lieu à l’existence d’autres espèces vivant dans d’autres milieux, en second lieu à la possibilité pour ces espèces d’émigrer vers la biocénose considérée, en troisième lieu à leur possibilité de s’insérer durablement dans le réseau trophique tout en y provoquant nécessairement une redistribution du flux de matière et d’énergie. En d’autres termes, l’enrichissement spécifique d’une biocénose dépend de ses rapports spatiaux avec d’autres communautés , sources éventuelles d’espèces différentes et de sa capacité à accueillir les populations immigrantes.
Les recherches sur ces phénomènes complexes ont été facilitées par l’étude des communautés insulaires qui fournissent d’excellents modèles pour aborder ces problèmes. C’est dans ce contexte que la théorie de Mac Arthur et Wilson, dite de l’équilibre dynamique , a été élaborée. Dans ses grandes lignes, elle postule que la richesse spécifique d’une communauté insulaire résulte d’un équilibre entre le flux d’immigration de nouvelles espèces (dépendant notamment de la distance de l’île aux biocénoses-sources) et du flux d’extinction des espèces s’installant dans l’île, flux qui dépend de la taille de celle-ci, de la variété de ses biotopes et de la plus ou moins grande ancienneté des communautés en place.
Dans l’ensemble, la théorie de Mac Arthur et Wilson a été souvent bien vérifiée, tant dans le cas d’îles vraies que d’îles «continentales», comme par exemple des sommets de montagne espacés. Elle s’applique cependant à une échelle de temps assez courte: elle ne prend pas en considération les processus de formation d’espèces nouvelles (spéciation) pouvant modifier le pool d’espèces potentiellement colonisatrices et elle ne tient compte qu’indirectement de l’ajustement interspécifique qui peut se produire au sein d’une biocénose (cf. infra : coévolution).
Progressivement approfondie grâce aux nombreux travaux qu’elle a suscités, la théorie de Mac Arthur et Wilson n’en constitue pas moins le cadre permettant de bien poser le problème de l’évolution des biocénoses: même si celles-ci ne sont pas insulaires au sens strict, l’insularisation, c’est-à-dire les phénomènes transitoires ou définitifs d’isolement spatial, joue un rôle considérable dans la formation des espèces et des communautés. C’est ainsi que la composition et la répartition actuelles des espèces dans les forêts équatoriales est la résultante de phases alternées de fragmentation et de réunification liées à la succession de périodes sèches et humides. En période sèche, les forêts se résolvent en «îles» séparées par des étendues herbacées. Les communautés de ces refuges forestiers évoluent alors indépendamment. Leur réunification en période pluvieuse met en présence des populations ayant parfois divergé au point d’avoir acquis l’isolement reproductif, mais ayant éventuellement continué d’utiliser les mêmes ressources. Ce sont alors de véritables espèces potentiellement compétitrices qui se trouvent en présence, ce qui doit conduire à un réajustement des peuplements dont elles font partie. Ce problème est de même nature que celui de l’insertion d’une espèce supplémentaire dans une biocénose insulaire.
Au-delà de ce problème de l’accroissement numérique des communautés se pose celui de l’ajustement fonctionnel des espèces au sein d’une biocénose. Les phénomènes de coévolution ont été surtout étudiés à propos de relations plantes-animaux ou proies-prédateurs: on a cherché à interpréter les interactions entre deux populations comme la résultante d’un processus évolutif dans lequel les modifications d’une population retentissent sur l’autre et réciproquement.
La multiplicité des relations interspécifiques existant au sein d’une biocénose, leur subtilité et leur spécificité souvent prononcées obligent à élargir cette perspective: dans une communauté où interfèrent relations trophiques, relations compétitrices et mutualisme, il est évident que toute transformation d’une population est susceptible de modifier ce réseau d’interactions et par conséquent les pressions sélectives auxquelles sont soumises les autres populations.
Il est classique de distinguer différents effets de la sélection naturelle. Elle peut être stabilisante lorsque un équilibre étant établi entre une population et son environnement, la sélection élimine les individus les moins adaptés à celui-ci. Avant que cet équilibre ne soit atteint, la sélection modifie progressivement la population jusqu’à ce qu’elle soit ajustée à son environnement: la sélection est alors directionnelle ou novatrice . Lorsque l’espèce est répartie entre des biocénoses isolées, soit par fragmentation d’une biocénose initiale, soit du fait de la colonisation d’autres biocénoses, les différentes populations ainsi formées peuvent subir des sélections directionnelles différentes et diverger peu à peu: globalement, la sélection naturelle est alors diversifiante.
Dans une biocénose en cours d’évolution, chaque population est ainsi soumise à une sélection directionnelle particulière mais, du fait des interactions entre espèces, bien que la sélection porte sur des pools géniques isolés, il y a détermination réciproque des pools géniques des populations interactives.
Le constat de la généralité de la coadaptation des espèces au sein des biocénoses conduit ainsi à admettre la généralité de la cosélection des populations et de leurs interactions. Lorsqu’une biocénose en voie de constitution accueille des populations supplémentaires, le jeu des interactions est modifié et la cosélection opère de façon directionnelle jusqu’à l’ajustement mutuel des populations: pour chacune, la sélection naturelle devient alors stabilisante.
Dans les conditions de l’équilibre dynamique, si le renouvellement est trop rapide (petite île proche des sources d’espèces immigrantes, par exemple), ce processus ne peut guère se développer. Dans les communautés trop isolées pour se renouveler fréquemment, en revanche, la coévolution peut aboutir à la formation d’espèces endémiques étroitement coadaptées.
Il semblerait que les biocénoses ayant atteint un niveau élevé de coadaptation se montrent résistantes à l’immigration de nouvelles populations. En fait, tout dépend de l’efficacité relative des stratégies adaptatives des espèces en place et des espèces colonisatrices: si celles-ci se montrent particulièrement aptes à exploiter le milieu, la compétition peut aboutir à l’élimination des espèces en place. Ce phénomène est à la base du renouvellement des flores et des faunes , processus maintes fois répété à l’échelle des temps géologiques.
L’enrichissement en espèces d’une biocénose ne peut donc avoir lieu que dans un contexte où l’introduction de populations nouvelles ne produit pas une perturbation importante du fonctionnement, mais constitue au contraire un facteur d’amélioration fonctionnelle de la communauté. Il peut y avoir par exemple élévation du rendement des transferts trophiques réalisés par une certaine guilde lorsque celle-ci s’enrichit en espèces, pour peu que le partage des ressources qui s’établit par coévolution conduise à une exploitation plus complète de celles-ci. Souvent, les espèces concernées sont issues de l’évolution divergente, dans des biocénoses séparées, de populations appartenant initialement à la même espèce. Elles ont alors fréquemment conservé des potentialités fonctionnelles voisines. L’accroissement de leur nombre dans une même biocénose peut alors accroître la capacité de stabilisation de celle-ci et ses potentialités d’adaptation aux changements du milieu.
Des biocénoses analogues, mais à richesses spécifiques différentes, peuvent ainsi avoir une plus ou moins grande efficience fonctionnelle et des stratégies adaptatives différentes. La stratégie cénotique d’une communauté est donc la conséquence de son histoire particulière, qui a été principalement déterminée par les contraintes du milieu et leurs variations à différentes échelles de temps, par les tentatives d’immigration d’espèces provenant d’autres biocénoses et enfin par les extinctions d’espèces qui provoquent à chaque fois un réajustement de la communauté.
Une telle approche évolutive doit permettre une interprétation cohérente de la variété des biocénoses. Elle ne peut guère être menée qu’à l’aide d’une analyse comparative de la structure et du fonctionnement de biocénoses analogues ayant évolué dans les contextes différents: l’écologie comparée , comme l’a permis à une autre échelle l’anatomie comparée, doit conduire à une meilleure compréhension de l’évolution.
5. Méthodologie écologique
Traits fondamentaux de la recherche
L’écologie a pour but premier de connaître le fonctionnement de populations et d’écosystèmes, c’est-à-dire d’ensembles d’éléments interactifs. Comme toutes les disciplines biologiques, l’écologie est donc une science fondamentalement systémique, tenue à d’incessants aller et retour entre l’analyse et la synthèse. Elle se particularise cependant du fait du niveau élevé de complexité des systèmes écologiques et surtout du fait que, sauf cas tout à fait exceptionnels, on ne peut espérer analyser ces systèmes dans leur totalité: on ne les connaît que par le biais d’échantillonnages et des représentations ne peuvent être construites que par combinaison et extrapolation des informations fournies par l’analyse des échantillons.
L’étude structurale et fonctionnelle des écosystèmes implique une approche quantitative visant notamment à déterminer des stocks et des flux de matière et d’énergie. Une telle quantification n’est évidemment possible que si l’on précise la nature des relations interspécifiques, grâce à une approche qualitative. À côté des relations d’exploitation, évidemment essentielles à connaître puisqu’elles définissent les voies de circulation de la matière et de l’énergie, les autres relations telles que compétition, symbiose, etc., sont également fondamentales, car toutes jouent un rôle, direct ou indirect, dans la détermination du débit de chaque transfert trophique.
L’écologie est par ailleurs confrontée à des difficultés de deux ordres qui, sans lui être propres, sont ici particulièrement importantes. Les unes sont dues à l’hétérogénéité spatiale des écosystèmes, les autres aux variations temporelles des facteurs abiotiques et des populations, variations qui ne sont pas toutes périodiques.
La complexité des biocénoses, la multiplicité et la variabilité des facteurs qui conditionnent leur fonctionnement, rendent ainsi les recherches écologiques particulièrement lourdes: seules des équipes nombreuses et diversifiées, dotées de moyens importants, peuvent les mener à bien. Ces moyens concernent à la fois les travaux de terrain et les études à mener en laboratoire; ils concernent aussi l’exploitation des données, la modélisation et la simulation des phénomènes: l’emploi de l’informatique, en effet, caractérise de plus en plus la recherche écologique.
Problèmes de quantification
Le fonctionnement d’un écosystème est sous la dépendance des facteurs abiotiques locaux dont il est indispensable de suivre les variations au cours du temps par des mesures qui posent des problèmes dont la solution relève de la physique et de la chimie. En revanche, la mesure des stocks de matière et d’énergie et la détermination des flux trophiques soulèvent des problèmes originaux. Chaque unité fonctionnelle d’une biocénose est une population: son étude quantitative suppose la mesure de son effectif, de la biomasse, des quantités correspondantes d’énergie, de carbone, d’azote, de phosphore, etc.; elle suppose aussi la mesure de la consommation, de la production et des dépenses d’entretien. Toutes les valeurs obtenues doivent être rapportées soit au volume de l’écosystème, soit à une unité de volume ou de surface, et bien entendu à une unité de temps. En général tout est rapporté à l’année, puisque le cycle fonctionnel d’un écosystème se ferme normalement avec le cycle saisonnier; la compréhension du fonctionnement oblige en outre à déterminer les variations des grandeurs étudiées, à l’intérieur du cycle annuel.
Le plus souvent, il n’est pas possible, pour des raisons pratiques, de mesurer de façon exhaustive l’effectif d’une population dans toute l’étendue de l’écosystème dont elle fait partie. On procède par prise d’échantillons correspondant à des volumes ou à des surfaces déterminées. La taille des échantillons et la méthode de dénombrement des individus doivent évidemment être adaptées à la population étudiée. Dans chaque cas, il importe d’apprécier la fidélité, la précision et la justesse de la technique envisagée. À la variété des organismes bactériens, végétaux et animaux qui composent une biocénose correspond une large panoplie de méthodes d’échantillonnage.
Du fait de l’hétérogénéité des écosystèmes, il est rare que la distribution spatiale des individus d’une population soit uniforme: leur densité varie d’un endroit à l’autre. De même, la structure démographique peut montrer des variations dans l’espace. En conséquence, la biomasse, la consommation, la production et les dépenses de maintenance varient aussi. Dans ces conditions, aucune mesure ponctuelle n’est suffisante pour assurer la représentativité d’un résultat. Cette difficulté est palliée par la répétition des échantillons dans l’espace, effectuée dans le but, d’une part, de dégager les caractéristiques moyennes de la population, d’autre part, de déterminer l’amplitude de variation des grandeurs mesurées.
L’utilisation des méthodes statistiques est indispensable non seulement pour traiter les données tirées de ces échantillons multiples, mais aussi – et d’abord – pour planifier les opérations de prélèvement.
La recherche des caractéristiques moyennes des populations se justifie dans le cadre de l’étude fonctionnelle globale d’un écosystème. En revanche, elle néglige les variations structurales dans l’espace dont l’étude à différentes échelles d’observation est pourtant tout aussi nécessaire, notamment pour comprendre l’organisation fonctionnelle des guildes composant l’écosystème et poser en termes corrects les problèmes de définition des niches. Ce sont encore des méthodes quantitatives qui sont utilisées, mais cette fois la planification des échantillonnages est conçue moins pour répondre aux exigences des méthodes statistiques que pour apporter une description qualitative précise de la structure spatiale de l’écosystème.
Analyse des interactions
La caractérisation globale d’un écosystème autorise à n’y distinguer que de grandes catégories fonctionnelles regroupant des populations à rôles trophiques analogues. Une telle approche synthétique doit être complétée par l’analyse des relations interspécifiques réelles et de l’influence des facteurs abiotiques sur chaque population: cette analyse relève donc à la fois de l’éthologie et de l’auto-écologie.
Les relations d’exploitation sont évidemment les premières à décrire puisqu’elles déterminent le réseau trophique. Selon les espèces, diverses méthodes peuvent être utilisées.
Chez les animaux, le comportement de prise de nourriture peut être observé dans la nature. Dans bien des cas, il est cependant plus efficace d’étudier le contenu du tractus digestif d’individus prélevés dans le milieu.
Lorsque la nourriture est broyée au point d’être méconnaissable, ou bien lorsque les animaux se nourrissent de liquides, tels les prédateurs à digestion externe, des méthodes immunologiques sont utilisables pour reconnaître dans les liquides absorbés des substances provenant d’une source supposée de nourriture.
Les relations mutualistes entre végétaux et animaux, ou entre animaux, peuvent être mises en évidence sur le terrain. En revanche, les relations de compétition sont particulièrement difficiles à déceler puisque, souvent, les distributions spatio-temporelles des organismes et leurs comportements sont coadaptés à la suite d’une évolution qui a eu justement pour conséquence une limitation sinon une suppression de la compétition.
Difficile à analyser avec précision chez les organismes supérieurs, les relations interspécifiques sont encore plus délicates à saisir lorsqu’elles mettent en jeu des micro-organismes, car l’identification de ceux qui sont réellement actifs dans un écosystème à un moment donné soulève des problèmes techniques qui ne sont pas encore surmontés.
Le double aspect du temps et ses conséquences méthodologiques
Les alternances du jour et de la nuit, la succession des saisons confèrent à la vie des écosystèmes un caractère périodique et le temps apparaît ainsi comme cyclique: à la même date, à la même heure, un écosystème se retrouve en principe dans le même état qu’un an plus tôt; comprendre son fonctionnement implique donc que les observations et les échantillonnages soient répétés de façon périodique.
En réalité, certains événements ne se reproduisent pas identiques à eux-mêmes d’un cycle à l’autre: c’est ainsi que, dans diverses régions, le climat montre souvent de considérables variations interannuelles, affectant par exemple les précipitations et leur répartition mensuelle. La dynamique de certaines populations peut s’en trouver modifiée, parfois immédiatement, parfois de façon différée. De ce fait, le temps est orienté et les populations suivent une histoire toujours unique qui peu à peu modèle leurs pools géniques: en toute rigueur, rien ne se répète jamais.
Bien entendu, à notre échelle d’observation, cela n’empêche pas de dégager pour chaque écosystème ses caractéristiques fonctionnelles moyennes: fruits d’une évolution souvent longue, elles ne présentent généralement pas de modifications profondes d’une année à l’autre. Il arrive cependant qu’une étude faite sur un seul cycle annuel fournisse des résultats trop particuliers pour être valablement représentatifs du passé comme du futur. Il est donc indispensable que certains écosystèmes fassent l’objet d’études à long terme pour que leurs changements éventuels puissent être observés et interprétés correctement.
Synthèse: modélisation et simulation
À partir des données collectées sur le terrain, il est possible de construire diverses représentations d’un écosystème, l’idéal étant de reproduire et de prévoir les variations temporelles de sa structure et de son fonctionnement. De ce point de vue, le développement de l’informatique a déjà permis des progrès considérables. Il est en effet devenu possible d’élaborer des modèles très complexes et d’en simuler le fonctionnement sur ordinateur.
Un modèle peut être conçu soit comme une représentation aussi correcte que possible du système, soit comme un moyen d’étudier ses propriétés et de tester la qualité des connaissances que l’on possède à son sujet. Dans les deux cas, l’élaboration du modèle passe par le choix des variables d’état représentant le système, celui des relations existant entre ces variables et des paramètres qui les caractérisent, celui enfin des facteurs externes conditionnant le fonctionnement du système (variables dites «de forçage»).
Une fois le modèle convenablement élaboré du point de vue mathématique, son fonctionnement peut être étudié par simulation sur ordinateur: on voit ainsi comment évoluent au cours du temps les variables d’état en fonction des variations imposées à l’entrée du modèle aux variables de forçage.
Des étapes importantes jalonnent la mise au point d’un modèle. On procède tout d’abord à une analyse de sensibilité qui permet de juger du comportement du modèle lorsque l’on modifie son état initial, certains paramètres ou même la conception de tel ou tel aspect de son fonctionnement, ou encore, plus simplement, lorsque l’on modifie les variables de forçage. Les appréciations portées sur la sensibilité dépendent des objectifs poursuivis, une très grande sensibilité pouvant être appréciable dans certains cas, préjudiciable dans d’autres.
La calibration intervient ensuite. Cette opération consiste à perfectionner peu à peu le modèle de façon que son fonctionnement se rapproche autant que possible de celui du système représenté. Ce dernier n’est évidemment connu qu’à partir de données recueillies pendant un temps limité, aussi rien ne dit qu’une représentation correctement calibrée sur la base de ces données se révélera capable de traduire convenablement le fonctionnement ultérieur du système étudié. Il se pose donc un problème de validité dont l’appréciation dépend pour une large part de la fonction assignée au modèle.
La modélisation et la simulation ne peuvent donc s’employer qu’avec de multiples précautions. Elles constituent néanmoins la meilleure manière de développer une réelle approche synthétique de la structure et du fonctionnement des systèmes écologiques, approche qui doit être soigneusement coordonnée avec les recherches menées sur le terrain afin de les orienter de façon efficace.
6. Applications de l’écologie
Exploitation de la production biologique
Bien qu’ils soient souvent en régression rapide, les écosystèmes naturels et semi-naturels jouent encore, au total, un rôle important. C’est le cas tout d’abord des écosystèmes aquatiques, qu’ils soient lacustres, fluviatiles et surtout marins, le cas aussi des écosystèmes forestiers, des garrigues et maquis, des savanes, des marécages et des déserts.
Dans les milieux aquatiques l’homme prélève quelques Algues, mais exerce surtout des activités de pêche, qui permettent un apport important de nourriture. Les progrès récents des techniques de détection et de capture ont été tels que l’exploitation dépasse bien souvent les possibilités de renouvellement des stocks. Dans le cas des Cétacés, chacun sait que le massacre a, malgré les règlements, dépassé largement les limites du raisonnable, mais la situation de certains Poissons, pour être moins spectaculaire, n’en est pas moins grave. Les problèmes écologiques liés à la pêche dans les milieux naturels dépassent à vrai dire souvent de simples questions de dynamique des populations: il touchent à l’équilibre de peuplements complexes vivant dans des milieux hétérogènes. L’urgence d’études globales de ces problèmes est de plus en plus grande, tant est rapide l’appauvrissement auquel a conduit une exploitation abusive et désordonnée. Le cas de nombreux secteurs des eaux littorales est typique à cet égard. La solution doit reposer sur des analyses poussées des possibilités réelles de production des divers milieux et des conditions d’une exploitation rationnelle, mais aussi, bien entendu, sur une réglementation stricte – et respectée – de l’intensité et des modalités de la pêche.
Sur les continents , l’exploitation par la chasse du monde animal autochtone a de nos jours perdu presque toute son importance pour l’alimentation de l’homme, sauf en quelques contrées reculées. Dans peu d’années, il n’y aura d’autre faune sauvage que celle spécialement protégée par l’homme et ce qui était du «gibier» ne proviendra plus que d’élevages, au même titre que le bétail domestique. De la même façon la cueillette de produits végétaux dans les milieux naturels a perdu pratiquement toute importance devant les cultures.
Les produits des forêts, en revanche, si celles-ci sont souvent en nette régression, notamment dans les pays tropicaux, garderont sans doute leur importance pour l’homme. On assistera probablement, dans les années à venir, à un emploi plus intensif de la production végétale non consommable. Il s’agit de récolter en plus grande abondance la matière végétale susceptible d’être employée comme source d’énergie («biomasse» des adeptes des énergies nouvelles). Outre la collecte plus complète du bois, petit et gros, dans les forêts actuellement exploitées, on veut mettre à profit les mauvais taillis, voire les maquis et garrigues. À l’utilisation directe comme combustible, on tend à ajouter la fabrication de combustibles liquides et gazeux – méthanol, éthanol et méthane – d’un emploi plus facile. Techniquement réalisables, ces opérations ne doivent toutefois pas faire illusion, car la production végétale reste limitée et par ailleurs liée à un apport de nutriments que fournit le retour au sol de la production. Une exploitation abusive risque ainsi d’entraîner rapidement une forte diminution de la production, sinon une stérilisation au moins temporaire des sols. Seules des études écologiques précises des stocks de nutriments et des modalités de leur renouvellement peuvent permettre de prévoir et donc de prévenir de telles impasses.
Artificialisation des écosystèmes
Depuis longtemps l’homme a cherché à accroître la production végétale dont il dispose et c’est ainsi que sont nées les cultures, devenues de nos jours des écosystèmes de plus en plus artificialisés. Nous n’insisterons pas sur les cultures, alimentaires ou industrielles, qui sont plus spécifiquement le domaine de l’agronomie, encore que leur gestion puisse sans doute gagner à tenir compte des données de l’écologie, mais les forêts elles-mêmes, dans nos régions tempérées au moins, sont exploitées de façon à produire au maximum du bois fort en général, parfois du bois de feu, et cela en favorisant certaines espèces. Le terme de «futaie jardinée» marque bien cette artificialisation. Un pas supplémentaire est franchi lorsque les forêts sont devenues des plantations artificielles d’arbres sélectionnés, appartenant souvent à des espèces exotiques, jardinées, traitées parfois. Ce sont de véritables cultures d’arbres. Les pâturages ont eux aussi, le plus souvent, été créés par l’homme et occupent l’emplacement d’antiques forêts, que ce soit dans les régions tempérées ou dans des régions tropicales humides. Au Mexique, en Amérique centrale, au Venezuela, par exemple, les pâturages sont même constitués d’herbes importées, originaires d’Afrique, dont le rendement est meilleur que celui des Graminées autochtones. Le problème est de maintenir de tels écosystèmes grâce à une gestion adéquate.
Non moins artificiel est évidemment le peuplement animal élevé dans ces pâturages, à dominance de bovins domestiques, l’élevage ayant remplacé la chasse et la faune sauvage ne représentant plus qu’une part infime de la consommation de l’humanité.
L’artificialisation de la production animale se retrouve même maintenant pour la faune aquatique avec l’extension des élevages de Poissons (truite, carpe...) et le développement de l’aquaculture marine jusqu’alors surtout limitée à certains coquillages (huîtres, moules). De tels élevages posent en fait à l’écologiste des problèmes innombrables et délicats de nutrition, d’aménagement du milieu, de sélection de races satisfaisantes, d’introduction d’espèces provenant d’autres régions, de lutte contre les parasites, d’associations favorables d’espèces animales ou végétales.
Un domaine plus récent, du moins à l’échelle industrielle, mais promis vraisemblablement à un grand avenir dans la lutte contre la faim, est celui de la culture du plancton, et notamment des Algues unicellulaires comme les Chlorelles. Sans doute utilisera-t-on aussi des cultures de Levures ou de Bactéries. On voit que, s’ils sont encore des «systèmes», certains écosystèmes peuvent devenir à ce point artificiels qu’ils n’ont plus guère de point commun avec les milieux naturels.
Quel qu’en soit le degré, l’artificialisation est toujours liée à deux caractéristiques inéluctables. La première est une consommation d’énergie qui peut atteindre des valeurs considérables et dépasser même parfois la production du système. C’est le cas évidemment des cultures et élevages purement artificiels, que justifie seule la qualité – la valeur marchande – du produit obtenu, mais c’est le cas aussi de certaines cultures en plein champ et l’on a depuis quelque temps attiré l’attention sur la dépense énergétique qu’exigeait par exemple la culture du maïs. La seconde caractéristique des écosystèmes artificialisés est leur instabilité: ils ne se maintiennent qu’à la faveur de soins constants, soins qui exigent parfois beaucoup d’énergie sous forme d’engrais, de travail mécanique, de pesticides... et qui risquent toujours d’être insuffisants pour maintenir un équilibre toujours précaire. Les exemples sont effectivement fréquents de cultures qui ont dû être abandonnées par suite d’une transformation irréversible du sol, ou d’attaques parasitaires excessives entraînées par la disparition des prédateurs habituels.
D’une manière très générale, c’est en vue de la recherche d’une productivité soutenue au cours du temps et d’un équilibre biologique non abusivement coûteux en énergie que l’écologiste doit faire entendre sa voix. Ainsi seront évitées des solutions techniques dont l’apparent intérêt immédiat masque les conséquences fâcheuses à long terme.
C’est dans cette optique que la lutte chimique par insecticides contre les parasites des cultures ne doit être employée qu’avec précaution. Si une lutte purement biologique faisant intervenir des ennemis naturels de ces parasites ne peut suffire, on doit tenter, par une étude écologique du problème qui fera notamment ressortir les périodes sensibles, d’associer à cette lutte biologique un emploi judicieux, limité dans le temps et l’espace, d’une quantité minimale d’insecticides. On a donné le nom de lutte intégrée à ces techniques, qui étendent considérablement le champ de l’écologie appliquée. Son efficacité peut être davantage encore accrue si les cultures elles-mêmes sont organisées en vue de cette lutte, au sein d’un plan global d’agriculture intégrée permettant, en quelque sorte, de prendre à contre-pied les cycles parasitaires grâce à des rotations adéquates des espèces végétales cultivées.
Lutte contre les pollutions
On appelle pollution toute intervention de l’homme dans les équilibres naturels par la mise en circulation de substances toxiques, nuisibles ou encombrantes, qui troublent ou empêchent l’évolution naturelle du milieu. La mise en évidence des sources de pollution et des moyens efficaces pour les combattre et les faire disparaître est devenue l’une des tâches essentielles de l’écologie appliquée. Les problèmes de pollution s’aggravent en effet de façon continuelle par le développement rapide et intensif des industries nucléaires, pétrolières et chimiques, par l’emploi sur une échelle de plus en plus vaste de substances désherbantes, insecticides et détergentes, enfin par des concentrations urbaines de plus en plus vastes et exigeantes.
La pollution radioactive est liée aux déchets des usines nucléaires et aux explosions atomiques; elle affecte tous les milieux terrestres, aquatiques et aériens. Certains organismes, qui ont la propriété de fixer électivement des isotopes radioactifs dans leurs tissus ou leur squelette, constituent des indicateurs écologiques du taux de radiocontamination de l’atmosphère ou de l’eau, mais aussi un danger par la concentration qu’ils opèrent. On sait les conséquences génétiques durables que peut avoir l’action mutagène des radiations sur les plantes et les animaux utiles, comme sur l’homme lui-même.
La pollution chimique affecte surtout l’air et l’eau mais, dans des zones très industrialisées, elle atteint même les sols. De nombreux produits chimiques peuvent être toxiques pour les plantes, les animaux et l’homme. Il y a d’abord ceux dont le but direct est d’intoxiquer certaines catégories d’organismes (mauvaises herbes, insectes nuisibles, rats), mais qui, déversés dans la nature en quantités excessives et incontrôlées, détruisent même des espèces utiles ou bouleversent leurs chaînes alimentaires (cas de l’arrosage massif et non sélectif de DDT, par exemple). Il y a aussi tous les sous-produits nuisibles de l’industrie. Parmi ceux-ci, le plus dangereux dans l’atmosphère est l’anhydride sulfureux, abondant dans les fumées industrielles; très souvent, de l’arsenic et d’autres produits toxiques mêlés à des poussières diverses se trouvent dans les fumées humides et noires (smog) qui, en l’absence de vent, forment de véritables dômes au-dessus des villes industrielles. Il y a même les engrais chimiques qui, employés trop souvent en quantités abusives, deviennent nuisibles, sinon dangereux. Qu’il suffise de citer les nitrates, très solubles, dont l’excès entraîne la pollution des eaux courantes et même des nappes souterraines. Enfin, une place à part, mais essentielle, est prise par les détergents chimiques, importante source de toxicité pour les peuplements des eaux douces.
La pollution organique concerne surtout les eaux. Le versement dans l’eau des déchets organiques (excréments, cadavres) est, lorsqu’il reste limité, une source essentielle de fertilité chimique des eaux continentales et, par leur intermédiaire, de la mer. En revanche, lorsque le niveau maximal que le milieu peut supporter est atteint, la démolition aérobie, accomplie en milieu oxygéné par toute une série d’organismes démolisseurs, des nécrophages aux bactéries, n’est plus possible. La flore bactérienne aérobie est alors supplantée par une flore anaérobie putréfiante, dont les produits métaboliques (hydrogène sulfuré, hydrocarbures, ammoniaque) sont toxiques pour les organismes aérobies, donc pour tous les animaux et les végétaux chlorophylliens.
La pollution due aux hydrocarbures , qui relève à la fois de la pollution chimique et de la pollution organique, est surtout importante en mer et sur les côtes. Les produits pétroliers sont un poison pour beaucoup d’organismes, une cause indirecte de mortalité pour bien d’autres (algues enduites de mazout, incapables de photosynthétiser, oiseaux aquatiques privés de leur enduit imperméable, petits Invertébrés englués), une présence désagréable pour l’homme et la cause de propriétés organoleptiques mauvaises pour les animaux filtrateurs, dont plusieurs (huîtres, moules) deviennent de véritables réservoirs de mazout.
La pollution par les détritus solides a pris beaucoup d’importance depuis la généralisation de l’emploi des matières plastiques polymères, pratiquement indestructibles, qui viennent encombrer le sol et les eaux autour de toutes les grandes agglomérations.
Dans tous les domaines, la lutte contre les pollutions passe par une connaissance approfondie du fonctionnement des écosystèmes... et plus encore, évidemment, par des précautions et des réglementations qui les limitent à la source!
Écologie et gestion du patrimoine naturel
Le développement des préoccupations relatives à la protection de la nature a conduit à concevoir de plus en plus celle-ci comme un patrimoine, héritage du passé et ressource pour le futur. Ce qui, pour beaucoup, était res nullius est ainsi perçu comme le bien de tous, notamment des générations à venir.
L’idée que la nature est pour l’homme une ressource aux multiples facettes n’est évidemment pas neuve: c’est le fait de la considérer comme patrimoine de la société qui ouvre de nouvelles perspectives. Désormais, les problèmes ne se posent plus seulement en termes de protection, mais aussi et plus encore en termes de gestion: les rapports des hommes avec la nature doivent être établis de telle sorte que les ressources offertes par celle-ci restent renouvelables. Cela ne concerne pas uniquement les milieux les plus sauvages, mais aussi tous ceux qui sont artificialisés à des degrés divers et dont certains sont maintenant abandonnés.
La notion de ressource naturelle est extrêmemement large: l’eau, une forêt, une zone humide, un paysage, une espèce sauvage, par exemple, sont autant de ressources effectives ou potentielles. Toute la difficulté d’une véritable politique de gestion tient à ce qu’elle doit non seulement assurer le renouvellement des ressources actuellement utilisées, mais aussi préserver l’avenir en évitant que ne soient compromises des resssources potentielles dont l’intérêt n’est pas encore perçu.
Dans ce contexte, l’écologie joue un rôle essentiel, d’une part en apportant le cadre théorique nécessaire à l’élaboration de saines politiques de gestion, d’autre part en fournissant les méthodes nécessaires à l’inventaire normalisé des ressources et à la caractérisation des conditions de leur renouvellement.
La biosphère est constituée d’écosystèmes faits de populations coadaptées. Tels qu’ils sont actuellement, ces écosystèmes résultent d’une histoire où l’homme est intervenu de façon plus ou moins sensible. Les propriétés présentes des populations et des biocénoses, fruits de cette histoire, définissent leurs stratégies adaptatives, c’est-à-dire leurs moyens d’évoluer. À partir de ces notions fondamentales, il est possible de donner son véritable sens au concept de conservation: la conservation de la nature n’est pas une opération de muséologie visant seulement à figer les produits du passé, mais c’est la mise en œuvre des moyens nécessaires au maintien ou même à l’amélioration des stratégies adaptatives des systèmes écologiques: conserver la nature, c’est lui conserver ses potentialités évolutives .
On comprend alors le rôle essentiel des recherches qui tentent de connaître ce qui, dans la structure et le fonctionnement des écosystèmes, détermine leurs potentialités adaptatives. Il est en particulier fondamental de préciser l’influence des dimensions des écosystèmes sur le maintien d’une richesse spécifique élevée et, pour chaque population, de pools géniques à forte richesse allélique. Cela est d’autant plus nécessaire que l’aménagement des territoires a le plus souvent comme conséquence le morcellement des écosystèmes: les fragments qui subsistent peuvent être trop réduits pour conserver toutes leurs potentialités adaptatives. La délimitation des réserves naturelles doit aussi tenir compte de considérations de ce type. D’une façon générale, l’organisation de l’espace doit ainsi s’appuyer sur les données de l’écologie.
Par ailleurs, l’écologie permet la constitution d’instruments de surveillance servant à contrôler l’évolution des écosystèmes et à déceler assez tôt l’instauration de dysfonctionnements. Les indicateurs biologiques sont des organismes ou des ensembles d’organismes qui traduisent d’une façon aussi directe et évidente que possible des modifications qualitatives et quantitatives de l’écosystème dont ils font partie. Lorsque ces modifications sont dues à des variations physico-chimiques du milieu, il pourrait être plus efficace de mesurer celles-ci directement. En fait, cela demande souvent des moyens coûteux, surtout lorsqu’il faut mettre au point des méthodes de détection spécifiques des substances nouvelles, éventuellement polluantes, que l’homme produit sans arrêt. Il est donc souvent plus commode de déceler les modifications du milieu à partir de leurs effets sur les êtres vivants, surtout si ceux-ci fournissent des indications nettes sur des phénomènes difficilement accessibles à la mesure. L’intérêt des indicateurs biologiques tient ainsi à plusieurs raisons. En premier lieu, certains organismes peuvent se comporter comme des amplificateurs exprimant par des changements marqués de petites variations des facteurs perturbant l’écosystème dont ils font partie, cela d’autant plus qu’il peut exister des phénomènes de seuil. En second lieu, les organismes peuvent se comporter comme des récepteurs cumulatifs (par exemple de polluants) ou comme des mémoires : des perturbations temporaires peuvent être mémorisées assez longtemps lorsqu’elles ont entraîné, dans une biocénose, des changements de structure importants. Ainsi, selon des modalités diverses, les organismes et les communautés qu’ils forment se comportent comme de véritables observatoires permanents de l’environnement dont les propriétés – amplification, cumulation, mémorisation – rendent possible le repérage de perturbations du milieu dont la détection directe pourrait être fort difficile.
En réalité, l’enjeu actuel n’est pas seulement de promouvoir une exploitation de la production biologique moins génératrice de pollution et mieux adaptée aux nécessaires restrictions d’énergie: c’est tout l’aménagement des territoires qui doit faire l’objet de choix rationnels. Ceux-ci auront à s’appuyer à la fois sur des monographies d’écosystèmes et sur l’étude des interactions entre ceux-ci, à l’échelle régionale.
Une telle démarche revient à fonder sur des bases biologiques solides une nouvelle géographie humaine, étudiant dans une perspective globale les rapports de l’homme avec la biosphère. Cette approche sera fructueuse dans la mesure où une interdisciplinarité véritable réalisera une étroite coopération de l’écologie et des sciences de l’homme.
écologie [ ekɔlɔʒi ] n. f.
• 1874, répandu v. 1968; du gr. oikos « maison » et -logie, d'apr. économie
1 ♦ Didact. Étude des milieux où vivent les êtres vivants ainsi que des rapports de ces êtres entre eux et avec le milieu (⇒ biocénose, bioclimatologie, biogéographie, biomasse, biosphère, biotope, écosystème, éthologie). Spécialiste de l'écologie. ⇒ écologiste, écologue. Écologie et protection de l'environnement, des cycles naturels.
2 ♦ Cour. Mouvement visant à un meilleur équilibre entre l'homme et son environnement naturel ainsi qu'à la protection de celui-ci. ⇒ écologisme. — Courant politique défendant ce mouvement.
● écologie nom féminin (allemand Ökologie) Science ayant pour objet les relations des êtres vivants (animaux, végétaux, micro-organismes) avec leur environnement, ainsi qu'avec les autres êtres vivants. Synonyme courant de écologisme. ● écologie (expressions) nom féminin (allemand Ökologie) Écologie culturelle, branche de l'anthropologie qui étudie les différences entre cultures en fonction de leur environnement physique.
écologie
n. f.
d1./d BIOL Science qui étudie les conditions d'existence d'un être vivant et les rapports qui s'établissent entre cet être et son environnement.
|| ANTHROP écologie culturelle: partie de l'anthropologie qui étudie les différences entre les cultures à partir des différences de l'environnement.
d2./d Cour. Protection de la nature, de l'environnement.
Encycl. L'écologie se subdivise en autoécologie (étude des rapports d'une seule espèce avec le milieu où elle vit), synécologie (étude des rapports des espèces appartenant à un même groupement avec le milieu où elles vivent) et dynamique des populations (modifications et causes de l'abondance des espèces dans un même milieu). Tous les problèmes relatifs au maintien des équilibres biologiques, à la conservation de la nature, à la protection des faunes et des flores, et à la survie du milieu naturel relèvent de l'écologie appliquée. L'écologie animale répond aux mêmes principes généraux. Elle étudie l'action des facteurs physiques (lumière, température, etc.) sur les animaux, mais aussi et surtout les innombrables interactions qui unissent ceux-ci entre eux ou aux végétaux. La notion de chaîne alimentaire, qui commence avec le plancton ou les algues et se termine avec les espèces carnivores, a incité les écologistes à étudier avec un intérêt particulier les milieux où les animaux ne peuvent survivre qu'au prix d'adaptations très perfectionnées (déserts, abysses, etc.). La protection de la nature (flore, faune, fleuves, océans) relève de l'écologie appliquée et des préoccupations relatives à l'environnement humain (villes, milieu rural, etc.).
⇒ÉCOLOGIE, subst. fém.
A.— BIOLOGIE
1. Science qui étudie les relations entre les êtres vivants (humains, animaux, végétaux) et le milieu organique ou inorganique dans lequel ils vivent. Le siècle se termine avec les grands traités (...) qui ouvrent les voies à l'écologie ou science de l'action du milieu sur les êtres vivants (Hist. gén. sc., t. 3, vol. 1, 1961, p. 440). L'écologie des protozoaires est déjà bien étudiée; le peuplement des divers biotopes (terre, mer, eau douce) et les adaptations morphologiques caractéristiques de chacun d'eux ont été analysés par de nombreux chercheurs (Hist. gén. sc., t. 3, vol. 2, 1964, p. 672:
• Connaître l'écologie des plantes fruitières présente un intérêt double :
— d'une part, cette science fournit d'utiles indications pour l'implantation des espèces et variétés en dehors de leurs centres originels. Ainsi, le pommier, originaire de régions élevées, croît particulièrement bien en altitude.
BOULAY, Arboric. et prod. fruit., 1961, p. 25.
2. P. ext. Étude des conditions d'existence et des comportements des êtres vivants en fonction de l'équilibre biologique et de la survie des espèces. On parle aussi parfois d'une biogéographie écologique ou écologie de la distribution géographique (Hist. gén. sc., t. 3, vol. 2, 1964, p. 682).
B.— P. anal., Sc. Soc. [En parlant de communautés humaines] Études des relations réciproques entre l'homme et son environnement moral, social, économique. Les communautés et sociétés humaines vivent dans un environnement auquel elles s'adaptent et qui réagit sur elles. L'étude de ce processus complexe qui met en jeu les systèmes technologiques et l'organisation sociale, est l'objet de l'écologie (GOLFIN 1972). Écologie criminelle (THINÈS-LEMP. 1975), écologie sociale (WILLEMS 1970).
Rem. En compos. et dans des créations récentes écologie se réduit au 1er élément éco- (gr. :  , « maison »); v. écotype; p. ext. écosystème. Ensemble des éléments naturels ou non composant l'environnement d'une espèce. Les écosystèmes peuvent être d'étendue très variable : une petite mare temporaire de quelques mètres carrés est un écosystème au même titre qu'une forêt de quelques centaines ou de quelques milliers d'hectares (P. AGUESSE, Clefs pour l'écologie, Paris, Seghers, 1971, p. 20).
, « maison »); v. écotype; p. ext. écosystème. Ensemble des éléments naturels ou non composant l'environnement d'une espèce. Les écosystèmes peuvent être d'étendue très variable : une petite mare temporaire de quelques mètres carrés est un écosystème au même titre qu'une forêt de quelques centaines ou de quelques milliers d'hectares (P. AGUESSE, Clefs pour l'écologie, Paris, Seghers, 1971, p. 20).
, « maison »); v. écotype; p. ext. écosystème. Ensemble des éléments naturels ou non composant l'environnement d'une espèce. Les écosystèmes peuvent être d'étendue très variable : une petite mare temporaire de quelques mètres carrés est un écosystème au même titre qu'une forêt de quelques centaines ou de quelques milliers d'hectares (P. AGUESSE, Clefs pour l'écologie, Paris, Seghers, 1971, p. 20).Prononc. :[ ]. Étymol. et Hist. 1910 (Actes du IIIe Congrès internat. de bot., Bruxelles, vol. 1, p. 120 [Cr. des travaux de Paul Jaccard et Charles Flahault] : Le terme « Écologie » comprend l'ensemble des relations existantes entre les individus végétaux ou les associations végétales d'une part et la station d'autre part (
]. Étymol. et Hist. 1910 (Actes du IIIe Congrès internat. de bot., Bruxelles, vol. 1, p. 120 [Cr. des travaux de Paul Jaccard et Charles Flahault] : Le terme « Écologie » comprend l'ensemble des relations existantes entre les individus végétaux ou les associations végétales d'une part et la station d'autre part ( = demeure = station = milieu). L'Écologie comprend l'étude des conditions de milieu et des adaptations des espèces végétales, soit prises isolément — Autécologie —, soit réunies en associations — Synécologie, ou étude des formations —). Empr. à l'all. Ökologie (composé du gr.
= demeure = station = milieu). L'Écologie comprend l'étude des conditions de milieu et des adaptations des espèces végétales, soit prises isolément — Autécologie —, soit réunies en associations — Synécologie, ou étude des formations —). Empr. à l'all. Ökologie (composé du gr.  « maison » et de
« maison » et de  « discours »), terme forgé en 1866 par le zoologiste et biologiste allemand E. H. Haeckel [1834-1919], relevé dans la préface de son ouvrage Natürliche Schöpfungsgeschichte 1867 (d'apr. Encyclop. brit., s.v. Haeckel); l'empr. à l'all. s'est peut-être fait par l'intermédiaire de l'angl. œcology (1873 tr. Haeckel's Hist. Creat. Pref. ds NED). Bbg. GIRAUD (J.), PAMART (P.), RIVERAIN (J.). Mots ds le vent. Vie Lang. 1970, pp. 328-329.
« discours »), terme forgé en 1866 par le zoologiste et biologiste allemand E. H. Haeckel [1834-1919], relevé dans la préface de son ouvrage Natürliche Schöpfungsgeschichte 1867 (d'apr. Encyclop. brit., s.v. Haeckel); l'empr. à l'all. s'est peut-être fait par l'intermédiaire de l'angl. œcology (1873 tr. Haeckel's Hist. Creat. Pref. ds NED). Bbg. GIRAUD (J.), PAMART (P.), RIVERAIN (J.). Mots ds le vent. Vie Lang. 1970, pp. 328-329.
]. Étymol. et Hist. 1910 (Actes du IIIe Congrès internat. de bot., Bruxelles, vol. 1, p. 120 [Cr. des travaux de Paul Jaccard et Charles Flahault] : Le terme « Écologie » comprend l'ensemble des relations existantes entre les individus végétaux ou les associations végétales d'une part et la station d'autre part ( = demeure = station = milieu). L'Écologie comprend l'étude des conditions de milieu et des adaptations des espèces végétales, soit prises isolément — Autécologie —, soit réunies en associations — Synécologie, ou étude des formations —). Empr. à l'all. Ökologie (composé du gr. « maison » et de « discours »), terme forgé en 1866 par le zoologiste et biologiste allemand E. H. Haeckel [1834-1919], relevé dans la préface de son ouvrage Natürliche Schöpfungsgeschichte 1867 (d'apr. Encyclop. brit., s.v. Haeckel); l'empr. à l'all. s'est peut-être fait par l'intermédiaire de l'angl. œcology (1873 tr. Haeckel's Hist. Creat. Pref. ds NED). Bbg. GIRAUD (J.), PAMART (P.), RIVERAIN (J.). Mots ds le vent. Vie Lang. 1970, pp. 328-329.
écologie [ekɔlɔʒi] n. f.
ÉTYM. 1874; all. Ecologie, 1866, Haeckel; didact. jusqu'en 1968-1970, où le mot s'est répandu; du grec oikos « maison, habitat », et -logie, d'après économie.
❖
1 Didact. Science qui étudie les milieux où vivent et se reproduisent les êtres vivants, ainsi que les rapports de ces êtres avec le milieu. ⇒ Bionomie, éthologie; autoécologie, synécologie; écographie, écophysiologie. || L'écologie étudie conjointement les relations des espèces entre elles et avec leur milieu, ainsi que la dynamique de leurs populations. ⇒ Écosystème (et biocénose, biotope); biosphère, écosphère; milieu. || Écologie quantitative, basée notamment sur l'évaluation de la production des écosystèmes (⇒ Biomasse). || Écologie animale (zooécologie), écologie végétale (phytoécologie). || Écologie éthologique. ⇒ Éco-éthologie. || Écologie humaine. || Écologie marine, écologie forestière. || Écologie d'une espèce, d'un milieu. || Spécialiste de l'écologie. ⇒ Écologiste (A., 1.), écologue. || Écologie et agriculture, écologie agricole (⇒ Agrobiologie). || Écologie et respect de l'environnement, des cycles naturels. || Utilisation de l'écologie dans la protection ou la restauration des équilibres naturels (aménagement du littoral, etc.).
1 L'écologie tend à combler le fossé que l'industrie a creusé entre l'homme et les animaux.
Emmanuel Berl, le Virage, p. 163.
♦ Écologie culturelle : étude des différences entre les cultures et civilisations humaines, en fonction de leur environnement.
♦ Par métonymie. Les réalités étudiées par l'écologie, les êtres vivants et leurs milieux. || Étudier l'écologie d'une zone.
2 (…) le Brésil cherche déjà sa défense et fait valoir non seulement les grandes difficultés du défrichement de l'Amazonie, mais le danger que présenterait, pour l'écologie mondiale, la disparition de forêts productrices d'oxygène.
A. Sauvy, Croissance zéro ?, p. 119.
➪ tableau Noms de sciences et d'activités à caractère scientifique.
2 Cour. Doctrine visant à une meilleure adaptation de l'homme à son environnement naturel, vivant (animaux, plantes) et non vivant, ainsi qu'à une protection de celui-ci (⇒ Écologisme); courant politique défendant cette doctrine. ⇒ Écolo (fam.), écologiste.
❖
DÉR. Écologique, écologisme, écologiste, écologue.
COMP. Autoécologie, phytoécologie, synécologie. — V. Éco-.
Encyclopédie Universelle. 2012.